遅延拡散型ニコルソン型ハエ方程式に対する大域的指数安定性の解析
A modified Nicholson’s blowflies equation accompanying distinct time-varying delays is established in this paper.
背景と学術的系譜
起源と学術的系譜
本稿で取り上げる問題は、数理生物学、特に個体群動態のモデリング分野に端を発する。歴史的には、ロジスティック方程式のような単純なモデルが個体群成長の記述に用いられてきた。しかし、現実世界の生物個体群は、生活環における時間遅延や空間移動のために、より複雑な挙動を示すことが多い。
ニコルソン型ハエ方程式の解析という特定の問題は、昆虫個体群のより現実的なモデルを作成しようとする研究者たちの間で浮上した。特に、成熟時間などの要因が個体数に大きく影響する。ハエは、その明確な生活段階と、遅延した成熟が個体数変動に及ぼす観察可能な影響のために、このような研究の古典的な対象となってきた。オリジナルのニコルソン型ハエ方程式は、遅延フィードバック機構を持つ個体群によく見られる振動的挙動を捉える遅延微分方程式である。
本稿は、空間拡散と複数の時間変動遅延を組み込んだ修正拡散型ニコルソン型ハエモデル(式1.1)を導入した参考文献[4]に代表される先行研究に基づいている。「拡散型」という側面は、成熟個体の空間的広がりを考慮し、「時間変動遅延」は、成熟期間が一定ではなく、環境要因によって時間とともに変化する可能性があることを反映している。
先行アプローチの根本的な限界または「ペインポイント」であり、本稿の主な動機は、古典的な力学系理論が、特定の条件下での正の定常状態の安定性を解析できないことにある。具体的には、[4]を含む以前の研究では、繁殖率が低い場合($\sum_{j=1}^m \frac{\beta_j}{\gamma} \leq 1$)のゼロ定常状態(すなわち、個体群の絶滅)の安定性を確立することに成功していた。しかし、繁殖率が高い場合($\sum_{j=1}^m \frac{\beta_j}{\gamma} > 1$)で、正の定常状態(安定した非ゼロの個体群レベル)が期待される場合、古典理論は失敗した。この失敗は、時間変動遅延を伴う非自律方程式の複雑な性質によるものであり、通常、従来の安定性解析に不可欠な「半フロー構造」を欠いていた。この理論的な障害は、[4]の結論部分で明確に未解決問題として特定されており、本稿まで未解決のままであった。著者らは、この限界を克服し、これらのより現実的で複雑なモデルにおける正の定常状態の大域的指数安定性を理解するための厳密な枠組みを提供するために、本稿を執筆する必要に迫られた。
直感的な領域用語
- ニコルソン型ハエ方程式: あるハエの群れの個体数が時間とともにどのように増減するかを記述する数学的な「物語」を想像してほしい。この物語には、新しいハエがどれだけ生まれ、どれだけ死に、そして最も重要なことに、幼虫が成虫になるまでどれくらいの時間がかかるかというルールが含まれている。これは、ハエの個体数の実際の増減を模倣するために設計された特定の公式である。
- 時間変動遅延: 種の種が花になるまでにかかる時間を考えてみてほしい。この「遅延」は常に同じではなく、暖かい晴れた日には短く、寒い曇りの日には長くなる可能性がある。ハエのモデルでは、「時間変動遅延」とは、ハエが成熟するまでにかかる時間が固定ではなく動的に変化することを意味し、モデルをより現実に忠実にする。
- 拡散型: コップ一杯の水に食紅を一滴垂らして広がる様子を想像してほしい。この文脈での「拡散型」とは、ハエが一箇所に留まるだけでなく、生息地全体を移動して広がることを意味する。この空間的な移動は、個体数が地域全体でどのように変化するかの重要な部分である。
- 大域的指数安定性: これは、自分で起き上がるおもちゃのようなものである。どこから始めようと、どのように押そうと、常に素早くスムーズに直立した安定した位置に戻る。 「大域的」とは、どの開始点からでも機能することを意味し、「指数関数的」とは、減衰の速い振動のように、その安定した位置に非常に効率的かつ迅速に落ち着き、平衡状態に達することを意味する。
記法表
| 記法 | 説明 |
|---|---|
| $\Theta(t,x)$ | 時間 $t$ および空間位置 $x$ におけるハエの個体群密度。 |
| $M$ | 滑らかな境界 $\partial M$ を持つ $\mathbb{R}^N$ の有界領域。 |
| $\Delta$ | ラプラシアン演算子、空間拡散を表す。 |
| $\gamma$ | 成熟ハエの単位時間あたりの死亡率($\gamma > 0$)。 |
| $m$ | それぞれ独自の遅延と密度依存パラメータを持つ、ハエの異なるグループの数。 |
| $\beta_j$ | グループ $j$ の単位時間あたりの産卵生産係数($\beta_j > 0$)。 |
| $r_j(t)$ | グループ $j$ の時間変動成熟遅延($r_j(t) > 0$)。 |
| $a_j$ | グループ $j$ の密度依存強度パラメータ($a_j > 0$)。 |
| $e^{-a_j \Theta(t - r_j(t), x)}$ | 未成熟段階における密度依存自己制限をモデル化する指数項。 |
| $\frac{\partial \Theta}{\partial n}(t,x) = 0$ | ノイマン境界条件、$\partial M$ の境界を横切るフラックスがないことを示す。 |
| $\psi(\theta, x)$ | 区間 $[-\tau, 0] \times M$ における個体群密度の初期履歴関数。ここで $\tau = \max_j \tau_j$ は最大遅延である。 |
| $X_+$ | 非負連続関数の空間、生物学的に現実的な個体群密度を表す。 |
| $\Theta^*$ | 方程式の唯一の正の定常状態解、安定した非ゼロの平衡個体群密度を表す。 |
| $A = \Delta - \gamma Id$ | ノイマン境界条件の下での $A$ の閉包によって生成される強連続半群、$Id$ は恒等演算子である。 |
| $T(t)$ | ノイマン境界条件の下での $A$ の閉包によって生成される強連続線形作用素半群。 |
| $\kappa^*$ | 大域的指数安定性の結果を述べる上で重要な $(0,1)$ の特定の定数。 |
| $\sum_{j=1}^m \frac{\beta_j}{\gamma}$ | 定常状態の存在と安定性に影響を与える、死亡率に対する総生産率を表す主要パラメータ。 |
| $\sum_{j=1}^m \frac{\beta_j}{a_j \gamma}$ | ゼロ定常状態の安定性に特に重要な別の主要パラメータ。 |
| $\sum_{j=1}^m \frac{\beta_j}{\gamma} e^{-a_j \Theta^*}$ | 定常状態の特性方程式から導出され、その安定性を決定するために使用される条件。 |
| $\lambda$ | 大域的指数安定性における指数的減衰率を表す正の定数。 |
| $N_0$ | 指数吸引性の初期上限を表す正の定数。 |
| $l, L$ | 摂動補題における解 $B(t)$ の下限と上限。 |
| $z^0$ | 解の下限の補題2.3の証明で使用される正の定数。 |
| $\delta_0$ | 補題2.4の証明で使用される小さな正の定数。 |
| $\xi$ | 解が特定の範囲内に束縛された後の時間閾値。 |
| $b^*$ | 時間 $\xi$ 後の解 $\Theta^0(t,x)$ の上限。 |
問題定義と制約
中核問題の定式化とジレンマ
本稿で取り上げる中核問題は、生物個体群モデルの長期的な挙動を理解することである。具体的には、著者らは、複数の時間変動遅延とノイマン境界条件を組み込んだ修正拡散型ニコルソン型ハエ方程式、すなわち初期境界値問題(IBVP)(1.1)を調査している。
出発点(入力/現在の状態)は、拡散と時間依存の成熟遅延などの要因を含めることで、現実世界の個体群動態をより正確に表現することを目指す、この複雑な数学モデルの存在である。先行研究、特に[4]は、特定の条件下(例:$\sum_{j=1}^m \frac{\beta_j}{\gamma} \le 1$ および $\sum_{j=1}^m \frac{\beta_j}{a_j \gamma} < 1$)でのゼロ定常状態解の安定性と大域的指数吸引性に関する基準を既に確立していた。これは、それらの条件下では個体群は最終的に絶滅することを意味する。
望ましい終点(出力/目標状態)は、IBVP (1.1)の正の定常状態解($\Theta^*$)が大域的指数安定性と吸引性を示す条件を決定することである。これは、個体群が非ゼロレベルで持続し安定化する可能性を理解するために重要である。本稿は、特にゼロ定常状態が不安定であることを示唆する条件である$\sum_{j=1}^m \frac{\beta_j}{a_j \gamma} > 1$ のシナリオを対象としている。これは、正の平衡状態の可能性を示唆する。
正確な欠落リンクまたは数学的ギャップは、時間変動遅延を伴う非自律方程式が含まれる場合に、この正の定常状態の安定性と指数吸引性を解析するための堅牢な理論的枠組みの欠如である。この特定の構成は、通常、古典的な力学系理論を適用するための前提条件である半フロー構造を欠いている。このギャップは、[4]の結論部分で未解決問題として明確に特定されており、この研究まで未解決のままであった。
研究者を閉じ込めてきた痛みを伴うトレードオフまたはジレンマは、生物学的現実性と数学的扱いやすさとの固有の対立である。複数の時間変動遅延($r_j(t)$)と異なる密度依存強度パラメータ($a_j$)を組み込むことは、ニコルソン型ハエ方程式を生物学的現象のより忠実な表現にする一方で、これらの特徴自体が重大な数学的複雑さを導入する。非自律性と時間変動遅延は半フロー構造を破壊し、特に正の定常状態に対する標準的な解析ツールを無効にする。したがって、モデルの記述能力を向上させることは、同時に、特に絶滅しない個体群に対する厳密な安定性解析を極めて困難にした。
制約と失敗モード
この問題は、主にモデルの数学的構造に起因する、著者らが直面したいくつかの厳しい現実的な壁のために、解決が非常に困難である。
- 半フロー構造の欠如(数学的制約): 最も重大な障害は、非自律方程式と時間変動遅延を備えたIBVP (1.1) が半フロー構造を持たないことである。これは、特に正の定常状態の安定性と吸引性を解析するために、多くの古典的な力学系理論を適用するための基本的な要件である。この構造がないと、位相空間解析や長期的な挙動予測のための標準的な技術は失敗する。
- 時間変動遅延(数学的制約): 複数の時間変動遅延 $r_j(t)$ を含めることは、定数遅延と比較して複雑さを増す。遅延自体が時間とともに変化し、システムの記憶と将来の状態に非定常な影響を与えるため、ダイナミクスは予測および解析がはるかに困難になる。これにより、システムは非自律的になり、リアプノフ汎関数やその他の安定性解析ツールの構築が複雑になる。
- 異なる密度依存強度パラメータ(数学的制約): モデルは、各遅延項に対して異なる密度依存強度パラメータ $a_j$ を許容する。この異質性(一部の先行研究[18]で仮定されているような、すべての $a_j$ が等しい均質な条件とは対照的)は、より多くの変動を導入し、一般的な安定性条件を見つけることをより困難にするため、解析をさらに複雑にする。
- 正の定常状態への焦点(数学的制約): 正の定常状態の大域的指数安定性を解析することは、ゼロ定常状態の解析よりも本質的に困難である。ゼロ定常状態は、線形化とより単純な解析を可能にすることが多いが、正の定常状態は、非自明な平衡状態の周りの非線形ダイナミクスを理解する必要があり、特に $\sum_{j=1}^m \frac{\beta_j}{a_j \gamma} > 1$ の条件下では、著しく複雑である。
- 理論的障害(理論的制約): 本稿で明確に述べられているように、非自律方程式と時間変動遅延の組み合わせは、「理論的障害」を生み出し、先行研究[4]では正の定常状態の安定性に関する問題が未解決のまま残されていた。既存の理論的枠組みは、これらの条件下での正の定常状態の大域的指数安定性を設定するには単純に不十分であった。著者らは、微分不等式のための新しい解析手法を開発し、摂動補題を適用して、これらの困難を首尾よく克服する必要があった。
なぜこのアプローチなのか
選択の必然性
ニコルソン型ハエのような方程式でモデル化される生物個体群の複雑なダイナミクスを調査する際、現実世界の複雑さを導入することは、しばしば従来の解析ツールの限界を押し広げる。本稿では、著者らは、標準的な手法が不十分であることが判明した重要な岐路に直面した。この認識の正確な瞬間は、問題の本質に由来する。すなわち、複数の時間変動遅延と異なる密度依存強度パラメータを組み込んだ遅延拡散型ニコルソン型ハエ方程式の正の定常状態の大域的指数安定性を解析することである。
著者らが3ページで述べているように、中心的な問題は、「時間変動遅延を伴う非自律方程式に関連するIBVP (1.1) は、通常、半フロー構造を欠いている」ことである。この半フロー構造の欠如は根本的な障害である。安定性解析の大部分の基礎を形成する古典的な力学系理論は、時間とともに解の進化を追跡するために、この構造に大きく依存している。それがなければ、これらの従来の「手法は、$\sum_{j=1}^m \frac{\beta_j}{\gamma} > 1$ の場合に、IBVP (1.1) の唯一の正の定常状態解の安定性と指数吸引性を解析することに失敗する。」この特定の条件は、死亡率でスケールされた出生率係数の合計が1を超える場合であり、以前は一部の著者自身の先行研究[4]でさえ未解決問題であった特に困難なシナリオを表す。
したがって、「新しい不等式技術、摂動補題、および線形作用素半群理論を組み合わせた」アプローチの選択は、単なる好みではなく、必然であった。これらの高度な解析ツールは、半フロー構造の単純化仮定なしに、システムの非自律性、時間変動性、および空間依存性を扱うことができる唯一の実行可能な解決策であった。
比較優位性
この解析アプローチの優位性は、計算速度やメモリフットプリントによって測定されるものではない。これは、アルゴリズム的なものではなく、理論的な数学論文であるためである。むしろ、その質的な利点は、以前は不可能であった、より現実的で複雑な生物学的モデルに対して厳密な解析的証明を提供できる能力にある。
構造的な利点は多岐にわたる。
1. 時間変動遅延の処理: 多くの初期の研究が遅延を定数に単純化していたのに対し、この方法は「数学的な観点から時間変動遅延関数に適している」(3ページ)。特に摂動補題は、遅延が固定されていないシステムを解析するための強力なツールであり、成熟時間や妊娠期間が変動する生物学的プロセスをより正確に表現することを可能にする。
2. 異なる密度依存パラメータの対処: 先行モデルでは、密度依存強度パラメータ ($a_j$) の均質性が仮定されることが多かった。本稿では、異なる $a_j$ 値を明示的に考慮しており、これは生物学的多様性をより正確に反映している。ここで開発された新しい不等式技術は、これらの変動パラメータによって導入される複雑さを管理するために正確に作成されており、このより一般的なシナリオで有効な安定性条件の導出を可能にする。
3. 未解決問題の解決: その優位性の最も説得力のある証拠は、「[4]からの未解決問題を解決した」(14ページ)ことである。これは、$\sum_{j=1}^m \frac{\beta_j}{\gamma} > 1$ の条件下で、正の定常状態の大域的指数吸引性を確立できなかった先行手法の限界を指す。現在の枠組みは、以前の「ゴールドスタンダード」手法が不十分であった場合に、包括的な解決策を提供する。
4. 一般化可能性: 著者らは、「解析的枠組みは、この特定のモデルに限定されるものではなく、開発された方法論は、密度依存パラメータを持つ他の遅延拡散型個体群モデルを研究するために容易に適合させることができる」(14ページ)と強調している。これは、直接的な範囲を超えて拡張される堅牢で一般化可能な理論的基盤を示唆している。
本質的に、この方法は、複数の時間変動遅延と異なる密度依存強度パラメータによってもたらされる技術的な困難を克服するための堅牢な数学的枠組みを提供し、新規かつより一般的な安定性基準につながるため、圧倒的に優れている。
制約との整合性
選択された解析方法は、問題の厳しい要件と完璧に「結婚」し、各制約に合わせた数学的ツールで対処している。
- 複数の時間変動遅延: 問題は、時間関数である遅延 $r_j(t)$ を明確に定義している。著者らの「解析と証明戦略は、時間変動遅延関数に適している」(3ページ)。摂動補題は、アプローチの重要な構成要素であり、このような遅延によって導入される非自律性を扱うために特別に設計されており、これらの動的な変化を考慮した安定性基準の導出を可能にする。
- 拡散性質とノイマン境界条件: ラプラシアン演算子 $\Delta \Theta(t,x)$ の存在は拡散を示し、ノイマン境界条件 $\frac{\partial \Theta}{\partial n}(t,x) = 0$ は孤立した生息地を意味する。線形作用素半群理論は、このような条件下での偏微分方程式(PDE)を解析するための理想的な数学的枠組みである。本稿では、$A = \Delta - \gamma Id$ を定義し、その閉包 $\bar{A}$ によって生成される強連続半群 $T(t)$ を利用している(4ページ)。この理論は、関数空間における解の存在、一意性、および特性を確立するために必要な機構を提供し、空間的文脈における安定性解析の基礎となる。
- 異なる密度依存強度パラメータ: モデルは、$a_j$ が異なることを許容しており、単純な均質な仮定からの逸脱である。注記3.2は、新しい証明方法が「複数の時間変動遅延と異なる密度依存強度パラメータによって引き起こされる技術的な困難を首尾よく克服した」と明確に述べている。「新しい不等式技術」は、これらの変動パラメータから生じる複雑さを管理するために正確に作成されており、このより一般的なシナリオで有効な安定性条件の導出を可能にする。
- 正の定常状態の大域的指数安定性: 最終目標は、大域的指数安定性を証明することである。新しい不等式技術、摂動補題、および線形作用素半群理論の組み合わせは、安定性だけでなく、定常状態への収束率を定量化する指数的安定性を確立するために必要な厳密な解析ツールを提供する。この包括的なアプローチにより、著者らは、特に $\sum_{j=1}^m \frac{\beta_j}{\gamma} > 1$ のような困難な条件下でも、正の定常状態に対してこの強力な安定性を保証する十分な基準を導出することができる。
代替案の却下
本稿は、この特定の問題に対して、他の一般的または伝統的なアプローチがなぜ失敗したのかを明確に説明している。主に暗黙的に却下されているのは、古典的な力学系理論である。
3ページで述べられているように、「時間変動遅延を伴う非自律方程式に関連するIBVP (1.1) は、通常、半フロー構造を欠いているため、古典的な力学系理論は、$\sum_{j=1}^m \frac{\beta_j}{\gamma} > 1$ の場合に、IBVP (1.1) の唯一の正の定常状態解の安定性と指数吸引性を解析することに失敗する。」これは直接的かつ明確な却下である。古典的な力学系理論は、アトラクタや安定性などの解の長期的な挙動を定義および解析するために、半フローの存在に依存している。時間変動遅延の導入はシステムを非自律的にし、この重要な半フロー構造を破壊し、多くの古典的なツールを適用不能にする。
さらに、著者らは、先行研究(自身の研究を含む)で使用された手法の十分性を暗黙的に却下している。注記3.2(11ページ)と結論(14ページ)は、「最近の研究[2, 3, 4, 7, 8, 16, 17]およびそこで引用されている参考文献は、異なる密度依存パラメータと複数の遅延を持つ拡散型ニコルソン型ハエモデルの正の定常状態の安定性と大域的指数吸引性に対処していない」と強調している。これは、これらの先行研究で採用された技術が、より単純なケース(例:定数遅延または均質なパラメータ)には価値があったとしても、本稿で提示されたより一般的で現実的なモデルには不十分であったことを示唆している。現在の研究は、これらの限界を具体的に対処し、より包括的で堅牢な解析的枠組みを提供している。
数学的・論理的メカニズム
マスター方程式
本稿の数学的エンジンの絶対的な核心は、ハエ個体群の時空間ダイナミクスを記述する修正拡散型ニコルソン型ハエ方程式である。これは、論文の式(1.1)として提示されている。
$$ \frac{\partial \Theta}{\partial t}(t, x) = \Delta \Theta(t,x) - \gamma \Theta(t, x) + \sum_{j=1}^{m} \beta_j \Theta(t - r_j(t), x) e^{-a_j \Theta(t - r_j(t), x)} \quad \text{in } (0, +\infty) \times M $$
この偏微分方程式は、ノイマン境界条件と初期履歴関数を伴う。
$$ \frac{\partial \Theta}{\partial x}(t, x) = 0 \quad \text{on } (0, +\infty) \times \partial M $$
$$ \Theta(\theta, x) = \psi(\theta, x) \geq 0, \quad (\theta, x) \in [-\tau, 0] \times M, \quad \psi \in X_+ $$
項ごとの解剖
マスター方程式の各構成要素を分解して、その数学的定義、物理的または論理的な役割、および選択された数学的操作の根拠を理解しよう。
-
$\frac{\partial \Theta}{\partial t}(t, x)$
- 数学的定義: これは、ハエ個体群密度 $\Theta$ の時間 $t$ に関する偏導関数である。特定の時間 $t$ および空間位置 $x$ における個体群密度の瞬間的な変化率を定量化する。
- 物理的/論理的役割: これは、個体群密度の純粋な変化率を表す。正の値は個体群の成長を示し、負の値は減少を示す。これは、方程式全体が記述しようとする従属変数である。
- なぜ偏導関数か: 個体群密度 $\Theta$ は時間 ($t$) と空間 ($x$) の両方で変化するため、空間分布を認識しながらその時間的進化を捉えるには偏導関数が不可欠である。
-
$\Delta \Theta(t,x)$
- 数学的定義: これは $\Theta(t,x)$ に適用されるラプラシアン演算子である。多次元空間領域では、各空間座標に関する $\Theta$ の二階偏導関数の合計である(例:2Dでは $\frac{\partial^2 \Theta}{\partial x_1^2} + \frac{\partial^2 \Theta}{\partial x_2^2}$)。これは、局所的な曲率、またはある点における $\Theta$ の値が無限小近傍の平均値からどれだけ異なるかを測定する。
- 物理的/論理的役割: これは拡散項である。成熟したハエの空間的な移動をモデル化する。ある点における個体群密度がその周囲よりも低い場合(正のラプラシアン)、個体はその点に移動する傾向があり、局所密度が増加する。逆に、密度が高い場合(負のラプラシアン)、個体は移動し、局所密度が減少する。この項は、個体群の空間的な広がりと均質化を考慮する。
- なぜ加算か: 拡散は個体群密度の全体的な変化に寄与する。拡散が増加を引き起こす場合は変化率に加算され、減少を引き起こす場合は減算される($\Delta \Theta$ は負になりうるため)。
-
$-\gamma \Theta(t, x)$
- 数学的定義: これは線形項であり、$\gamma > 0$ は定数係数で、現在の個体群密度 $\Theta(t,x)$ に掛けられている。
- 物理的/論理的役割: これは死亡率項を表す。$\gamma$ は成熟ハエの単位時間あたりの死亡率である。負の符号は、この項が常に個体群密度を減少させることを示している。これは、既存の個体群の一部が時間とともに死滅するという自然な減衰プロセスをモデル化する。
- なぜ減算か: 死亡率は個体を個体群から直接除去するため、変化率から減算される。
-
$\sum_{j=1}^{m} \beta_j \Theta(t - r_j(t), x) e^{-a_j \Theta(t - r_j(t), x)}$
- 数学的定義: これは $m$ 個の異なる項の合計であり、それぞれがハエの特定のグループ $j$ からの出生寄与を表す。各項は、出生率係数 $\beta_j$、遅延時間における個体群密度 $\Theta(t - r_j(t), x)$、および指数因子 $e^{-a_j \Theta(t - r_j(t), x)}$ の積である。
- 物理的/論理的役割: これは出生率項であり、ニコルソン型ハエモデルの最も複雑で特徴的な部分である。
- $\beta_j$: これはグループ $j$ の単位時間あたりの産卵生産係数である。潜在的な子孫の数をスケーリングする正の定数である。
- $\Theta(t - r_j(t), x)$: これは、過去の時間 $t - r_j(t)$ における個体群密度を表す。項 $r_j(t) > 0$ は、グループ $j$ の時間変動成熟遅延である。この遅延は、時間 $t$ における新しい成熟ハエ(出生)が、 $r_j(t)$ 時間単位前に存在した未成熟個体に由来するため、重要である。$r_j(t)$ の時間変動性は、成熟期間に対する環境影響などを反映する動的な生物学的プロセスを反映している。
- $e^{-a_j \Theta(t - r_j(t), x)}$: この指数項は、未成熟段階における密度依存自己制限または死亡率を導入する。$a_j > 0$ は、グループ $j$ の密度依存強度パラメータである。遅延時間における個体群密度 $\Theta(t - r_j(t), x)$ が増加すると、この指数項は減少し、実効出生率を抑制する。これは、未成熟ハエの間の資源競争、過密、または捕食/病気の増加などの現象をモデル化し、成虫への生存率を低下させる。 $xe^{-ax}$(ここで $x = \Theta(\cdot)$)という特定の形式は、ニコルソン型モデルの特徴であり、出生率が個体群の増加とともに最初は増加するが、非常に高い密度では低下することを示唆している。
- 合計 $\sum_{j=1}^{m}$: モデルは $m$ 個の異なるハエのグループを考慮しており、それぞれが潜在的に独自の出生パラメータ($\beta_j$, $r_j(t)$, $a_j$)を持つ。合計は、これらのグループすべてからの総出生寄与を集計する。
- なぜ加算か: 出生は個体数を増加させるため、出生寄与の合計全体が変化率に加えられる。
- 項内のなぜ乗算か: 出生率は、潜在的な産卵数($\beta_j \Theta(\cdot)$)と成虫への生存確率($e^{-a_j \Theta(\cdot)}$)の積である。これらの要因は乗算的に組み合わされて、新しい成虫の実際数を決定する。
- なぜ指数関数か: 指数形式は、個体群動態、特にニコルソン型ハエ方程式における密度依存効果をモデル化する標準的な方法であり、個体群密度に対する非単調な出生率応答を捉える。
-
境界条件: $\frac{\partial \Theta}{\partial x}(t, x) = 0 \quad \text{on } (0, +\infty) \times \partial M$
- 数学的定義: これはノイマン境界条件であり、空間変数 $x$ に関する $\Theta$ の法線方向の導関数が領域 $M$ の境界 $\partial M$ 上でゼロであることを示している。
- 物理的/論理的役割: この条件は、境界を横切るフラックスなしを意味する。生物学的には、生息地 $M$ が孤立しており、領域へのまたは領域からのハエの純粋な移動がないことを意味する。これは、閉鎖生態系における一般的な仮定である。
- なぜゼロとの等価性か: ゼロの法線方向の導関数は、境界を横切る純粋な流れまたは交換がないことを意味する。
-
初期条件: $\Theta(\theta, x) = \psi(\theta, x) \geq 0, \quad (\theta, x) \in [-\tau, 0] \times M, \quad \psi \in X_+$
- 数学的定義: 個体群密度 $\Theta$ は、最大遅延区間 $[-\tau, 0]$ 内のすべての過去の時間 $\theta$ と空間領域 $M$ 全体にわたる初期関数 $\psi(\theta, x)$ によって定義される。関数 $\psi$ は非負でなければならない。
- 物理的/論理的役割: これは個体群の初期履歴を設定する。方程式が時間変動遅延 $r_j(t)$ を含むため、時間 $t$ における個体群の将来の進化は、様々な過去の時間 $t - r_j(t)$ における個体群の状態に依存する。したがって、システムは計算を開始するために、過去のある期間(最大遅延 $\tau$ まで)の個体群密度の履歴を知る必要がある。非負制約($\psi \geq 0$)は生物学的に不可欠であり、個体群密度は負になりえないためである。
- なぜ区間上の関数か: 遅延が存在するということは、システムの現在の状態が過去の状態の影響を受けることを意味する。これを正確にモデル化するには、初期条件は、関連する遅延期間全体にわたる個体群の履歴を提供する必要がある。
ステップバイステップの流れ
この数学的エンジンを通過するハエ個体群密度を表す抽象的なデータポイントのライフサイクルを追ってみよう。これは、個体群動態のための連続的な組み立てラインと考えることができる。
- 入力: 現在の状態と履歴: 任意の瞬間 $t$ および空間位置 $x$ において、システムは現在の個体群密度 $\Theta(t,x)$ を取り込む。重要なのは、遅延のため、履歴の個体群密度 $\Theta(s,x)$ ($s \in [t-\tau, t]$) にもアクセスすることである。この履歴データは、$t \leq 0$ の場合の初期条件 $\psi$ によって提供される。
- 死亡率計算: 現在の個体群密度 $\Theta(t,x)$ は、直ちに「死亡率ユニット」に供給される。ここでは、死亡率 $\gamma$ が掛けられ、この積 $-\gamma \Theta(t,x)$ は、その瞬間に死亡する個体数を示す。これは個体群への直接的な減少である。
- 拡散処理: 同時に、現在の個体群密度 $\Theta(t,x)$ とその近傍の空間は「拡散ユニット」に供給される。ラプラシアン演算子 $\Delta$ は純粋な空間フラックスを計算する。周囲の領域の密度が高い場合、個体は流入し、$\Theta(t,x)$ を増加させる。$\Theta(t,x)$ が高い場合、個体は流出し、それを減少させる。このユニットは空間的な混合を保証する。
- 遅延出生生産(並列処理): $m$ 個の異なるハエグループそれぞれについて、「出生生産ユニット」が並列で動作する。
- それは時間 $t - r_j(t)$ (グループ $j$ の時間変動遅延によって決定される特定の過去の瞬間)まで遡り、その履歴時点から個体群密度 $\Theta(t - r_j(t), x)$ を取得する。
- この履歴密度は、グループの単位あたり産卵生産係数 $\beta_j$ で乗算される。
- 次に、この潜在的な出生率は「密度依存フィルター」を通過する。指数項 $e^{-a_j \Theta(t - r_j(t), x)}$ は出生率を調整する。履歴個体群が非常に高かった場合、このフィルターは実効出生率を大幅に低下させ、成熟中の資源不足や競争をモデル化する。
- 各グループの出生生産ユニットの出力は、新しい成虫へのそのグループの特定の寄与である。
- 集計と純変化: 死亡率ユニット、拡散ユニット、およびすべての $m$ 個の出生生産ユニットからの出力は、「合計ユニット」に供給される。このユニットは、これらの寄与をすべて合計して、個体群密度の総瞬間変化率 $\frac{\partial \Theta}{\partial t}(t, x)$ を計算する。
- 時間積分: この計算された変化率は、$\Theta(t,x)$ の連続的な進化を駆動する。システムは、この率を時間とともに積分し、次の無限小瞬間の個体群密度を更新する。
- 境界強制: このプロセス全体を通じて、「境界制御メカニズム」は、ノイマン境界条件が厳密に維持されることを保証する。領域の端を横切る個体をもたらす拡散運動はすべて防止され、個体群は $M$ 内に閉じ込められる。
この連続的なフィードバックループは、現在および過去の状態が将来の変化を決定し、モデルがハエ個体群の時空間における複雑なダイナミクスをシミュレートすることを可能にする。
最適化ダイナミクス
本稿の焦点は、典型的な機械学習の意味でのパラメータの「最適化」や損失関数の最小化ではない。むしろ、ここでいう「最適化ダイナミクス」とは、システムが安定な平衡状態に向かって進化するというシステムの固有の挙動を指す。中核的な目的は、正の定常状態 $\Theta^*$ の大域的指数安定性を証明することである。
- 定常状態をアトラクタとして: 定常状態 $\Theta^*$ は、個体群密度が時間とともに変化しなくなる構成であり、$\frac{\partial \Theta}{\partial t}(t, x) = 0$ を意味する。空間的に均質な場合、これは $\Theta^*$ が $-\gamma\Theta^* + \sum_{j=1}^{m} \beta_j\Theta^*e^{-a_j\Theta^*} = 0$ を満たすことを意味する。解析の目的は、この $\Theta^*$ がシステム位相空間における強力なアトラクタとして機能することを示すことである。
-
大域的指数安定性: これは望ましい「最適化」結果である。
- 安定性: システムが $\Theta^*$ の近くから開始した場合、近くに留まり、最終的に $\Theta^*$ に戻る。
- 大域的吸引性: 初期正の個体群分布 $\psi$(生物学的に現実的である限り、すなわち非負)に関わらず、システムは最終的に $\Theta^*$ に収束する。これは、$\Theta^*$ がすべての可能な初期条件の究極の運命であることを意味する。
- 指数関数的: $\Theta^*$ への収束は漸近的であるだけでなく、指数関数的な速度で発生する。現在の個体群 $\Theta(t,x)$ と定常状態 $\Theta^*(x)$ との差は、通常、正の定数 $C$ と $\lambda$ を用いて $|\Theta(t,x) - \Theta^*(x)| \leq C e^{-\lambda t}$ と表され、指数関数的に速く減衰する。これは、システムがどれだけ速く「落ち着く」かを定量化する。
-
収束のメカニズム(解析ツール): 本稿では、反復アルゴリズムではなく、この収束を実証するために洗練された解析ツールのセットを採用している。
- 微分不等式と比較原理: 著者らは、方程式の解の上限と下限を構築する。実際の解がこれらの上限と下限の間に「挟まれている」ことを示し、これらの上限と下限自体が $\Theta^*$ に収束することを示すことで、解の収束を証明する。これは、微分方程式の長期的な挙動を解析するための基本的なツールである。
- 摂動補題: この補題は、遅延微分方程式の解の有界性を確立するために使用される。有界性は、安定性を証明するための重要な前提条件であり、個体群密度が無限に増加したりゼロに崩壊したりしないことを保証する(ゼロ定常状態の場合を除く)。
- 線形作用素半群理論: 拡散項 $\Delta \Theta(t,x) - \gamma \Theta(t,x)$ は、線形作用素 $A = \Delta - \gamma Id$ によって生成されると見なすことができる。ノイマン境界条件の下でのこのような作用素によって生成される強連続半群の理論は、基盤となる線形ダイナミクスを理解するための枠組みを提供する。半群 $T(t)$ のコンパクト性や強正性などの特性は、完全な非線形システムの挙動を解析するために活用される。
- リアプノフ様解析: 明示的にリアプノフ関数として名前が付けられていない場合でも、証明ではしばしば補助関数(または汎関数)を構築し、その時間微分が負であることが示される。これは、システムの「エネルギー」または定常状態からの「距離」が継続的に減少していることを意味し、それを $\Theta^*$ に駆動する。例えば、補題2.4は、関連する常微分方程式の指数吸引性を直接証明しており、これはリアプノフ安定性理論から導出される古典的な結果である。
- 逐次法/数学的帰納法: 時間変動遅延のため、解はしばしば successive time intervals (例:$[0, \sigma]$, $[\sigma, 2\sigma]$, etc.)にわたって構築および解析される。この方法は、著者らが解の局所的な特性を大域的なものに拡張することを可能にし、安定性の証明を時間領域全体にわたって構築する。
- 平均値定理と特定の不等式: 標準的な数学ツールが、正確な上限を導出し、指数的減衰を示すために適用される。例えば、不等式 $|\text{fe}^{-f} - \text{ge}^{-g}| \leq \frac{1}{e} |\text{f} - \text{g}|$ は、定常状態からの偏差への非線形出生項の寄与を制御するために使用される。
本質的に、「最適化ダイナミクス」は、システムをその唯一の正の定常状態に駆動する微分方程式内の固有の力である。数学的解析は、これらの力が大域的かつ指数関数的な収束を保証するのに十分に強いことを厳密に証明し、システムの位相空間を、$\Theta^*$ が堅牢で大域的に吸引的な平衡状態となるように形成する。本稿で導出された条件(例:$\sum \beta_j/\gamma$ のパラメータ)は、この安定した挙動が保証されるパラメータを定義する。
結果、限界、結論
実験デザインとベースライン
理論的発見、特に大域的指数安定性に関する定理3.1を厳密に検証するために、著者らは遅延拡散型ニコルソン型ハエ方程式の特定のインスタンス、すなわちIBVP (4.1) に焦点を当てた数値シミュレーションを設計した。これは、他のモデルとの直接的な数値競争ではなく、新しい解析的枠組みが、以前は対処されていなかった複雑さを処理できることを示す重要なデモンストレーションであった。
設定は、一次元空間領域 $M = [-1, 1]$ を含み、3つの異なる時間変動遅延、$r_1(t) = e^{-\sin^2 t}$, $r_2(t) = 2e^{-\sin^2 t}$, $r_3(t) = 4e^{-\sin^2 t}$ を組み込んだ。重要なのは、密度依存強度パラメータ $a_j$ も異なる値 ($a_1 = 1, a_2 = 101/100, a_3 = 103/100$) に選択されたことであり、これは先行文献でしばしば見られる均質性の仮定に違反する。単位あたり死亡率 $\gamma$ は3に設定され、出生率係数 $\beta_j$ はすべて $e$ に設定された。シミュレーションの初期値関数は0.6に設定された。
この文脈での「敗者」は、数値的に打ち負かされた特定のベースラインモデルではなく、むしろ既存の理論的枠組みの限界そのものであった。本稿は、先行研究(例:[7, 8, 9, 10, 11, 16, 17, 19])が、時間変動遅延と、そのような複雑な条件下での正の定常状態の安定性/指数吸引性を考慮できなかったことを明確に述べている。したがって、実験のデザインは、著者らが開発した新しい解析手法が、以前のアプローチが理論的に不十分であったり、問題が未解決のまま残されていたりした状況で、実際に解決策を提供できることを示すことであった。数値例は、これらの困難で、以前は解決不可能であった条件下での、本稿自身の理論的結果の正確さの直接的な検証として機能した。
証拠が証明するもの
数値シミュレーションは、著者らが提案した中核的な数学的メカニズムが、少なくとも選択された特定のパラメータに対しては、実際に機能したという決定的な、否定できない証拠を提供した。複雑で時間変動する遅延と異なる密度依存強度パラメータを持つIBVP (4.1) をシミュレートすることにより、著者らは正の定常状態 $\Theta^* \approx 0.848$ を計算した。重要なステップは、大域的指数安定性、特に補題2.4の条件(2.13)の条件がこれらのパラメータによって満たされていることを確認することであった。簡単な計算により、$\sum_{j=1}^m \frac{\beta_j}{\gamma} e^{-a_j \Theta^*} \approx 1.5252$ であり、これは確かに1より大きく、$\sum_{j=1}^m \frac{\beta_j}{a_j \gamma} e^{-a_j \Theta^*} \approx 0.886883$ であり、これは1より小さいことが確認された。これらの計算は、$\kappa^* \approx 0.432857$ とともに、定理3.1の理論的前提条件が満たされていることを確認した。
図4.1に示されている視覚的な証拠は、IBVP (4.1) の解が時間とともに計算された正の定常状態 $\Theta^* \approx 0.848$ に漸近的に収束することを示している。初期値0.6から開始して、個体群密度は進化し、この安定した平衡状態に落ち着き、大域的漸近安定性を示している。この視覚的な確認は、基盤となる数学的条件の成功した検証と相まって、新しい不等式技術、摂動補題、および線形作用素半群理論を含む著者らの新しい解析手法が、以前は未解決問題であったシナリオで大域的指数安定性の十分な基準を実際に提供したことを、徹底的に証明した。それは、彼らの枠組みが複数の時間変動遅延と異なる密度依存強度パラメータを実際に処理でき、それによって先行研究を拡張および改善したことを示した。
限界と将来の方向性
本稿は大きな進歩を遂げているが、その限界を認識し、将来の発展の方向性を考慮することが重要である。
一つの直接的な限界は、説得力があるにもかかわらず、数値例が特定のパラメータセットと一次元空間領域に限定されていることである。それは複雑なケースの理論的枠組みを検証するが、完全なパラメータ空間や高次元シナリオを探索するものではない。ベースラインモデルの「敗北」は理論的なものであり(そのような問題を解析できないこと)、パフォーマンスや精度の直接的な数値比較ではなかった。
著者らによって明確に強調されている重要な未解決の質問は、IBVP (1.1) の大域的ダイナミクスを、特に条件 $\sum_{j=1}^m \frac{\beta_j}{\gamma} e^{-a_j \Theta^*} > 1$ の下で、複数の時間変動遅延とともに特徴づけることに関するものである。現在の研究は、ゼロ定常状態の安定性または正の定常状態の特定の範囲内の場合、この合計が1未満の場合に焦点を当てている。この条件が逆転した場合や確立された範囲外の場合のダイナミクスを探索することは、振動や分岐のようなより複雑な挙動を明らかにする可能性のある、重要な次のステップとなるだろう。
将来を見据えると、いくつかの多様な視点がさらなる研究を刺激する可能性がある。
- より広範なシステムへの一般化: 著者らは、彼らの解析的枠組みが他の遅延拡散型個体群モデル、例えばフィッシャー-KPP方程式やマッキー・グラスシステムに適合可能であると示唆している。将来の研究では、この方法論をこれらのモデルや他の関連する生態学的または生物学的モデルに適用し、同様の複雑な遅延およびパラメータ条件下での安定性特性を確立することで、この適応性を明示的に実証できるだろう。
- 空間的不均一性と境界条件の影響: 現在の研究では、ノイマン境界条件を持つ有界空間領域を使用しており、孤立した生息地を意味する。異なる境界条件(例:境界での固定個体群のためのディリクレ条件、または周期条件)や、より複雑で不均一な空間構造の影響を調査することで、生息地の接続性と環境の変動性が安定性にどのように影響するかを明らかにできるだろう。
- 確率性と環境ノイズ: 現実世界の生物システムは、ほとんど決定的ではない。遅延拡散型ニコルソン型ハエ方程式に確率的摂動または環境ノイズを組み込むことは、現実性をさらに高めるだろう。そのようなノイズがグローバル指数安定性にどのように影響するか、あるいは確率共鳴やノイズ誘発遷移のような現象を引き起こすかどうかを解析することは、魅力的で実践的に関連性のある方向性となるだろう。
- 数値的手法と計算効率: これらのモデルの複雑さが増すにつれて、それらのシミュレーションの計算コストも増加する。時間変動遅延と異なるパラメータを持つ遅延反応拡散方程式に特別に調整された、より効率的で堅牢な数値スキームを開発することは、より大きなシステム、より長い時間スケール、およびより高い次元の探索を可能にし、理論的進歩を補完するだろう。
- 生物学的解釈と予測能力: 数学的厳密性を超えて、これらの安定性条件の生物学的意味合いをさらに深く掘り下げる必要がある。遅延パラメータや密度依存強度の特定の範囲は、ハエや他の種の個体群動態の観察とどのように相関するのか?このモデルは、個体群の発生、絶滅、または害虫管理における制御戦略の効果について、検証可能な予測を行うために使用できるのか?これは、抽象的な数学と応用生態学の間のギャップを埋めるだろう。
- クリティカルパラメータの特定: 実用的な応用のためには、安定性を支配する最も影響力のあるパラメータを特定することが鍵となる。感度分析を使用して、どのパラメータ(例:特定の遅延、出生率、死亡率、または密度依存強度)が大域的指数安定性に最も大きな影響を与えるかを理解できる。これは、生物学における実験デザインや保全活動に情報を提供する可能性がある。
これらの点を対処することにより、将来の研究は、遅延拡散システムの数学的理解を拡張するだけでなく、様々な科学技術分野における予測ツールとしての有用性を高めることができるだろう。
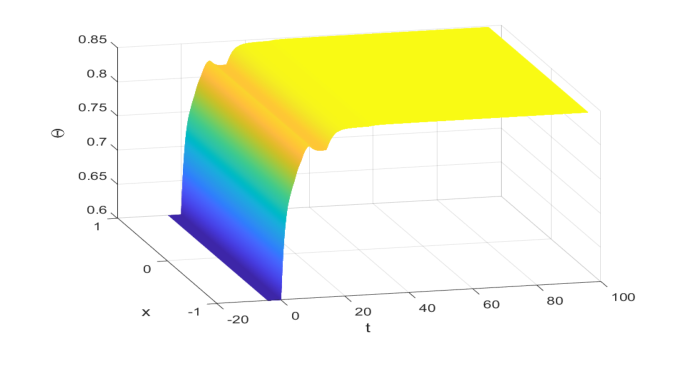 Figure 4. 1: The global asymptotic stability of Θ∗≈0.848 to (4.1) with initial value function 0.6
Figure 4. 1: The global asymptotic stability of Θ∗≈0.848 to (4.1) with initial value function 0.6