Глобальный экспоненциальный анализ устойчивости для диффузионного уравнения Николсона для мясных мух с запаздыванием
Проблема, рассматриваемая в данной статье, берет свое начало в области математической биологии, в частности, в моделировании динамики популяций.
Предыстория и академическая родословная
Происхождение и академическая родословная
Проблема, рассматриваемая в данной статье, берет свое начало в области математической биологии, в частности, в моделировании динамики популяций. Исторически для описания роста популяций использовались простые модели, такие как логистическое уравнение. Однако реальные биологические популяции часто демонстрируют более сложное поведение, особенно из-за временных запаздываний в жизненных циклах и пространственного перемещения.
Конкретная проблема анализа уравнения Николсона для мясных мух возникла, когда исследователи стремились создать более реалистичные модели популяций насекомых, где такие факторы, как время созревания, значительно влияют на численность популяции. Мясные мухи, в частности, стали классическим объектом таких исследований благодаря своим четко выраженным стадиям развития и наблюдаемому влиянию запаздывающего созревания на колебания численности популяции. Оригинальное уравнение Николсона для мясных мух является дифференциальным уравнением с запаздыванием, которое отражает колебательное поведение, часто наблюдаемое в популяциях с механизмами обратной связи с запаздыванием.
Данная статья основывается на предыдущих работах, в частности на ссылке [4], где была представлена модифицированная диффузионная модель Николсона для мясных мух (Уравнение 1.1), включающая как пространственную диффузию, так и множественные изменяющиеся во времени запаздывания. "Диффузионный" аспект учитывает пространственное распространение взрослых особей, в то время как "изменяющиеся во времени запаздывания" отражают тот факт, что периоды созревания могут быть не постоянными, а изменяться со временем из-за факторов окружающей среды.
Фундаментальным ограничением или "болевой точкой" предыдущих подходов, и основной мотивацией для данной статьи, является неспособность классической теории динамических систем анализировать устойчивость положительного стационарного состояния при определенных условиях. В частности, в предыдущих работах, включая [4], была успешно установлена устойчивость нулевого стационарного состояния (т.е. вымирание популяции) при низкой скорости воспроизводства ($\sum_{j=1}^m \frac{\beta_j}{\gamma} \leq 1$). Однако при высокой скорости воспроизводства ($\sum_{j=1}^m \frac{\beta_j}{\gamma} > 1$), и, следовательно, при ожидаемом наличии положительного стационарного состояния (стабильный, ненулевой уровень популяции), классическая теория оказалась неэффективной. Эта неудача была вызвана сложной природой неавтономных уравнений с изменяющимися во времени запаздываниями, которые обычно не обладают "структурой полупотока", являющейся критически важной для традиционного анализа устойчивости. Это теоретическое препятствие было явно идентифицировано как открытая проблема в заключительных замечаниях [4] и оставалось нерешенным до появления данной статьи. Авторы были вынуждены написать эту статью, чтобы преодолеть это ограничение и предоставить строгую основу для понимания глобальной экспоненциальной устойчивости положительного стационарного состояния в этих более реалистичных и сложных моделях.
Интуитивные термины предметной области
- Уравнение Николсона для мясных мух: Представьте себе математическую "историю", описывающую, как группа мясных мух растет и сокращается со временем. Эта история включает правила того, сколько новых мух рождается, сколько умирает, и, что крайне важно, сколько времени требуется детенышу мухи, чтобы стать взрослой особью. Это конкретная формула, разработанная для имитации реальных взлетов и падений популяции мясных мух.
- Изменяющиеся во времени запаздывания: Подумайте о времени, которое требуется семени, чтобы вырасти в цветок. Это "запаздывание" не всегда одинаково; оно может быть короче в теплую, солнечную погоду и длиннее в холодную, пасмурную. В модели мясных мух "изменяющиеся во времени запаздывания" означают, что время, необходимое мясной мухе для созревания, не фиксировано, а динамически меняется, делая модель более соответствующей действительности.
- Диффузионный: Представьте каплю пищевого красителя, распространяющуюся в стакане воды. "Диффузионный" в данном контексте означает, что мясные мухи не остаются на одном месте; они перемещаются и распространяются по своей среде обитания. Это пространственное движение является важной частью того, как их популяция меняется на определенной территории.
- Глобальная экспоненциальная устойчивость: Это похоже на самовосстанавливающуюся игрушку. Независимо от того, как вы ее толкнете или откуда начнете, она всегда быстро и плавно вернется в свое вертикальное, устойчивое положение. "Глобальная" означает, что это работает из любой начальной точки, а "экспоненциальная" означает, что она очень эффективно и быстро приходит в это устойчивое положение, подобно быстро затухающим колебаниям, достигая равновесия.
Таблица обозначений
| Обозначение | Описание |
|---|---|
| $\Theta(t,x)$ | Плотность популяции мясных мух во времени $t$ и пространственной локации $x$. |
| $M$ | Ограниченная пространственная область в $\mathbb{R}^N$ с гладкой границей $\partial M$. |
| $\Delta$ | Оператор Лапласа, представляющий пространственную диффузию. |
| $\gamma$ | Суточная смертность взрослых мясных мух на одну особь ($\gamma > 0$). |
| $m$ | Количество различных групп мясных мух, каждая со своими запаздываниями и параметрами, зависящими от плотности. |
| $\beta_j$ | Коэффициент производства потомства в сутки на одну особь для группы $j$ ($\beta_j > 0$). |
| $r_j(t)$ | Изменяющееся во времени запаздывание созревания для группы $j$ ($r_j(t) > 0$). |
| $a_j$ | Параметр интенсивности, зависящий от плотности, для группы $j$ ($a_j > 0$). |
| $e^{-a_j \Theta(t - r_j(t), x)}$ | Экспоненциальный член, моделирующий самоограничение, зависящее от плотности, на стадии развития молоди. |
| $\frac{\partial \Theta}{\partial n}(t,x) = 0$ | Граничное условие Неймана, указывающее на отсутствие потока через границу $\partial M$. |
| $\psi(\theta, x)$ | Начальная функция истории для плотности популяции в интервале $[-\tau, 0] \times M$, где $\tau = \max_j \tau_j$ — максимальное запаздывание. |
| $X_+$ | Пространство неотрицательных непрерывных функций, представляющих биологически реалистичные плотности популяции. |
| $\Theta^*$ | Единственное положительное стационарное решение уравнения, представляющее собой стабильную, ненулевую равновесную плотность популяции. |
| $A = \Delta - \gamma Id$ | Линейный оператор, представляющий диффузию и смертность, где $Id$ — единичный оператор. |
| $T(t)$ | Сильно непрерывная полугруппа линейных операторов, порожденная замыканием $A$ при граничных условиях Неймана. |
| $\kappa^*$ | Определенная константа в $(0,1)$, критически важная для формулировки результатов глобальной экспоненциальной устойчивости. |
| $\sum_{j=1}^m \frac{\beta_j}{\gamma}$ | Ключевой параметр, представляющий общее репродуктивное число относительно смертности, влияющее на существование и устойчивость стационарных состояний. |
| $\sum_{j=1}^m \frac{\beta_j}{a_j \gamma}$ | Еще один ключевой параметр, особенно актуальный для устойчивости нулевого стационарного состояния. |
| $\sum_{j=1}^m \frac{\beta_j}{\gamma} e^{-a_j \Theta^*}$ | Условие, выведенное из характеристического уравнения стационарного состояния, используемое для определения его устойчивости. |
| $\lambda$ | Положительная константа, представляющая скорость экспоненциального затухания в глобальной экспоненциальной устойчивости. |
| $N_0$ | Положительная константа, представляющая начальную границу для экспоненциальной аттрактивности. |
| $l, L$ | Нижняя и верхняя границы для решения $B(t)$ в лемме о флуктуациях. |
| $z^0$ | Положительная константа, используемая в доказательстве Леммы 2.3 для нижних границ решений. |
| $\delta_0$ | Малая положительная константа, используемая в доказательстве Леммы 2.4. |
| $\xi$ | Временной порог, после которого решения ограничены в определенном диапазоне. |
| $b^*$ | Верхняя граница для решения $\Theta^0(t,x)$ после времени $\xi$. |
Определение проблемы и ограничения
Основная постановка проблемы и дилемма
Основная проблема, рассматриваемая в данной статье, заключается в понимании долгосрочного поведения модели биологической популяции. В частности, авторы исследуют модифицированное диффузионное уравнение Николсона для мясных мух, представленное как начальная краевая задача (НПЗ) (1.1), которая включает множественные изменяющиеся во времени запаздывания и граничное условие Неймана.
Отправная точка (Входное/Текущее состояние) — существование этой сложной математической модели, призванной более точно отражать реальную динамику популяций, включая такие факторы, как диффузия и зависящие от времени запаздывания созревания. Предыдущие исследования, в частности [4], уже установили критерии устойчивости и глобальной экспоненциальной аттрактивности нулевого стационарного решения для этой модели при определенных условиях (например, $\sum_{j=1}^m \frac{\beta_j}{\gamma} \le 1$ и $\sum_{j=1}^m \frac{\beta_j}{a_j \gamma} < 1$). Это означает, что при этих условиях популяция в конечном итоге вымирала.
Желаемое конечное состояние (Выходное/Целевое состояние) — определить условия, при которых положительное стационарное решение ($\Theta^*$) НПЗ (1.1) демонстрирует глобальную экспоненциальную устойчивость и аттрактивность. Это имеет решающее значение для понимания того, как популяция может сохраняться и стабилизироваться на ненулевом уровне. Статья конкретно нацелена на сценарий, когда $\sum_{j=1}^m \frac{\beta_j}{a_j \gamma} > 1$, условие, при котором нулевое стационарное состояние неустойчиво, что подразумевает возможность положительного равновесия.
Точное недостающее звено или математический пробел — отсутствие надежной теоретической основы для анализа устойчивости и экспоненциальной аттрактивности этого положительного стационарного состояния, когда модель включает неавтономные уравнения с изменяющимися во времени запаздываниями. Эта конкретная конфигурация обычно не обладает структурой полупотока, которая является предпосылкой для применения классической теории динамических систем. Этот пробел был явно идентифицирован как открытая проблема в заключительных замечаниях [4] и оставался нерешенным до данной работы.
Болезненный компромисс или дилемма, которая поставила в тупик предыдущих исследователей, — это присущий конфликт между биологической реалистичностью и математической решаемостью. В то время как включение множественных изменяющихся во времени запаздываний ($r_j(t)$) и различных параметров интенсивности, зависящих от плотности ($a_j$), делает уравнение Николсона для мясных мух более точным представлением биологических явлений, эти самые особенности создают значительные математические сложности. Неавтономный характер и изменяющиеся во времени запаздывания нарушают структуру полупотока, делая стандартные аналитические инструменты неэффективными для положительного стационарного состояния. Таким образом, улучшение описательной силы модели одновременно сделало ее строгий анализ устойчивости, особенно для невымирающей популяции, чрезвычайно трудным.
Ограничения и режимы отказа
Эта проблема чрезвычайно сложна для решения из-за нескольких жестких, реалистичных стен, с которыми столкнулись авторы, в основном из-за математической структуры модели:
- Отсутствие структуры полупотока (Математическое ограничение): Самым значительным препятствием является то, что НПЗ (1.1) с неавтономными уравнениями и изменяющимися во времени запаздываниями не обладает структурой полупотока. Это фундаментальное требование для применения многих классических теорий динамических систем для анализа устойчивости и аттрактивности, особенно для положительных стационарных состояний. Без этой структуры стандартные методы анализа фазового пространства и прогнозирования долгосрочного поведения не работают.
- Изменяющиеся во времени запаздывания (Математическое ограничение): Включение множественных изменяющихся во времени запаздываний, $r_j(t)$, вместо постоянных запаздываний, добавляет уровень сложности. Динамику становится гораздо труднее предсказывать и анализировать, поскольку само запаздывание меняется со временем, влияя на память системы и ее будущее состояние непостоянным образом. Это делает систему неавтономной и усложняет построение функций Ляпунова или других инструментов анализа устойчивости.
- Различные параметры интенсивности, зависящие от плотности (Математическое ограничение): Модель допускает различные параметры интенсивности, зависящие от плотности, $a_j$, для каждого члена запаздывания. Эта гетерогенность (в отличие от однородного условия, когда все $a_j$ равны, как предполагалось в некоторых предыдущих работах, например [18]) еще больше усложняет анализ, внося больше вариативности и затрудняя поиск общих условий устойчивости.
- Фокус на положительном стационарном состоянии (Математическое ограничение): Анализ глобальной экспоненциальной устойчивости положительного стационарного состояния по своей сути более сложен, чем нулевого стационарного состояния. Нулевое стационарное состояние часто позволяет линеаризацию и более простой анализ, но положительное стационарное состояние требует понимания нелинейной динамики вокруг нетривиального равновесия, что значительно сложнее, особенно при условии $\sum_{j=1}^m \frac{\beta_j}{a_j \gamma} > 1$.
- Теоретическое препятствие (Теоретическое ограничение): Как явно указано в статье, комбинация неавтономных уравнений и изменяющихся во времени запаздываний создала "теоретическое препятствие", которое оставило проблему устойчивости положительного стационарного состояния нерешенной в предыдущих исследованиях [4]. Существующие теоретические рамки были просто недостаточны для установления глобальной экспоненциальной устойчивости положительного стационарного состояния при этих условиях. Авторам пришлось разработать новые аналитические методы для дифференциальных неравенств и использовать лемму о флуктуациях для успешного преодоления этих трудностей.
Почему такой подход
Неизбежность выбора
При изучении сложной динамики биологических популяций, особенно тех, что моделируются такими уравнениями, как уравнение Николсона для мясных мух, введение реальных сложностей часто раздвигает границы традиционных аналитических инструментов. В данной статье авторы столкнулись с критическим моментом, когда стандартные методы оказались недостаточными. Точный момент осознания этой проблемы проистекает из самой природы задачи: анализ глобальной экспоненциальной устойчивости диффузионного уравнения Николсона для мясных мух с запаздыванием, включающего множественные изменяющиеся во времени запаздывания и различные параметры интенсивности, зависящие от плотности.
Основная проблема, как сформулировали авторы на странице 3, заключается в том, что "НПЗ (1.1), связанная с неавтономными уравнениями с изменяющимися во времени запаздываниями, обычно не обладает структурой полупотока". Это отсутствие структуры полупотока является фундаментальным препятствием. Классическая теория динамических систем, которая составляет основу для большей части анализа устойчивости, сильно полагается на эту структуру для отслеживания эволюции решений во времени. Без нее эти обычные методы просто "не могут анализировать устойчивость и экспоненциальную аттрактивность для уникального положительного стационарного решения НПЗ (1.1) при $\sum_{j=1}^m \frac{\beta_j}{\gamma} > 1$". Это конкретное условие, когда сумма коэффициентов рождаемости, масштабированная коэффициентом смертности, превышает единицу, представляет собой особенно сложный сценарий, который ранее был открытой проблемой, даже в более ранних работах некоторых из тех же авторов [4].
Следовательно, выбор использования "новых методов неравенств в сочетании с леммой о флуктуациях и теорией полугрупп линейных операторов" был не просто предпочтением, а необходимостью. Эти передовые аналитические инструменты были единственным жизнеспособным решением, способным справиться с неавтономным, изменяющимся во времени и пространственно-зависимым характером системы без упрощающего предположения о структуре полупотока.
Сравнительное превосходство
Превосходство данного аналитического подхода измеряется не скоростью вычислений или потреблением памяти, поскольку это теоретическая математическая статья, а не алгоритмическая. Вместо этого его качественное преимущество заключается в его способности предоставлять строгие аналитические доказательства для более реалистичной и сложной биологической модели, чем это было возможно ранее.
Структурное преимущество многогранно:
1. Обработка изменяющихся во времени запаздываний: В отличие от многих более ранних исследований, которые упрощают запаздывания до постоянных, этот метод "хорошо подходит для функций запаздывания, изменяющихся во времени, с математической точки зрения" (стр. 3). Лемма о флуктуациях, в частности, является мощным инструментом для анализа систем, где запаздывания не фиксированы, позволяя более точно отражать биологические процессы, где время созревания или периоды беременности могут колебаться.
2. Решение проблем, связанных с различными параметрами, зависящими от плотности: Предыдущие модели часто предполагали однородность параметров интенсивности, зависящих от плотности ($a_j$). Данная статья явно рассматривает различные значения $a_j$, что является более точным отражением биологического разнообразия. Новые методы неравенств, разработанные здесь, имеют решающее значение для управления дополнительной сложностью, вносимой этими изменяющимися параметрами.
3. Решение открытой проблемы: Наиболее убедительным доказательством его превосходства является его успех в "решении открытой проблемы из [4]" (стр. 14). Это относится к неспособности предыдущих методов установить глобальную экспоненциальную аттрактивность положительного стационарного состояния при условии $\sum_{j=1}^m \frac{\beta_j}{\gamma} > 1$. Текущая структура предоставляет комплексное решение, где предыдущие "золотые стандарты" методов оказались недостаточными.
4. Обобщаемость: Авторы подчеркивают, что их "аналитическая структура не ограничивается данной конкретной моделью, и разработанная методология может быть легко адаптирована для изучения других диффузионных популяционных моделей с запаздыванием и параметрами, зависящими от плотности" (стр. 14). Это указывает на надежную и обобщаемую теоретическую основу, выходящую за рамки непосредственной сферы действия уравнения Николсона для мясных мух.
По сути, этот метод является подавляюще превосходящим, поскольку он предоставляет надежную математическую основу для преодоления технических трудностей, создаваемых множественными изменяющимися во времени запаздываниями и различными параметрами, зависящими от плотности, что приводит к новым и более общим критериям устойчивости.
Соответствие ограничениям
Выбранный аналитический метод идеально "сочетается" с жесткими требованиями задачи, решая каждое ограничение с помощью специализированных математических инструментов:
- Множественные изменяющиеся во времени запаздывания: Проблема явно определяет запаздывания $r_j(t)$, которые являются функциями времени. "Анализ и стратегия доказательства авторов хорошо подходят для функций запаздывания, изменяющихся во времени" (стр. 3). Лемма о флуктуациях, ключевой компонент их подхода, специально разработана для обработки неавтономного характера, вносимого такими запаздываниями, позволяя выводить критерии устойчивости, учитывающие эти динамические изменения.
- Диффузионный характер и граничное условие Неймана: Наличие оператора Лапласа $\Delta \Theta(t,x)$ означает диффузию, а граничное условие Неймана $\frac{\partial \Theta}{\partial n}(t,x) = 0$ подразумевает изолированную среду обитания. Теория полугрупп линейных операторов является идеальной математической основой для анализа дифференциальных уравнений в частных производных (ДУЧП) при таких условиях. В статье определяется $A = \Delta - \gamma Id$ и используется сильно непрерывная полугруппа $T(t)$, порожденная ее замыканием $\bar{A}$ (стр. 4). Эта теория предоставляет необходимый аппарат для установления существования, единственности и свойств решений в функциональных пространствах, что является фундаментальным для анализа устойчивости в пространственном контексте.
- Различные параметры интенсивности, зависящие от плотности: Модель допускает, что $a_j$ могут быть различными, что отличается от более простых однородных предположений. Замечание 3.2 (стр. 11) явно указывает, что новый метод доказательства "успешно преодолел технические трудности, вызванные множественными изменяющимися во времени запаздываниями и различными параметрами интенсивности, зависящими от плотности". "Новые методы неравенств" точно разработаны для управления сложностями, возникающими из-за этих изменяющихся параметров, что позволяет выводить условия устойчивости, действительные для этого более общего сценария.
- Глобальная экспоненциальная устойчивость для положительного стационарного состояния: Конечная цель — доказать глобальную экспоненциальную устойчивость. Комбинация новых методов неравенств, леммы о флуктуациях и теории полугрупп линейных операторов предоставляет строгие аналитические инструменты, необходимые для установления не только устойчивости, но и экспоненциальной устойчивости, которая количественно определяет скорость сходимости к стационарному состоянию. Этот комплексный подход позволяет авторам вывести достаточное условие, гарантирующее эту сильную форму устойчивости для положительного стационарного состояния, даже в сложных условиях, таких как $\sum_{j=1}^m \frac{\beta_j}{\gamma} > 1$.
Отклонение альтернатив
В статье четко излагается, почему другие популярные или традиционные подходы потерпели бы неудачу для данной конкретной проблемы. Основная альтернатива, косвенно отвергнутая, — это классическая теория динамических систем.
Как указано на странице 3, "поскольку НПЗ (1.1), связанная с неавтономными уравнениями с изменяющимися во времени запаздываниями, обычно не обладает структурой полупотока, классическая теория динамических систем не может анализировать устойчивость и экспоненциальную аттрактивность для уникального положительного стационарного решения НПЗ (1.1) при $\sum_{j=1}^m \frac{\beta_j}{\gamma} > 1$". Это прямое и недвусмысленное отвержение. Классическая теория динамических систем полагается на существование полупотока для определения и анализа долгосрочного поведения решений, такого как аттракторы и устойчивость. Введение изменяющихся во времени запаздываний делает систему неавтономной, нарушая эту критически важную структуру полупотока и делая многие классические инструменты неприменимыми.
Более того, авторы косвенно отвергают достаточность методов, использованных в предыдущих работах, включая некоторые из своих собственных. Замечание 3.2 (стр. 11) и заключение (стр. 14) подчеркивают, что "недавние исследования [2, 3, 4, 7, 8, 16, 17] и цитируемые в них ссылки не касались устойчивости и глобальной экспоненциальной аттрактивности положительного стационарного состояния для диффузионной модели Николсона для мясных мух с различными параметрами, зависящими от плотности, и множественными запаздываниями". Это подразумевает, что методы, использованные в этих более ранних работах, хотя и ценные для более простых случаев (например, постоянные запаздывания или однородные параметры), были недостаточны для более обобщенной и реалистичной модели, представленной в данной статье. Текущая работа конкретно устраняет эти ограничения, предоставляя более комплексную и надежную аналитическую основу.
Математический и логический механизм
Основное уравнение
Абсолютным ядром математического аппарата данной статьи является модифицированное диффузионное уравнение Николсона для мясных мух, которое описывает пространственно-временную динамику плотности популяции мясных мух. Оно представлено как уравнение (1.1) в статье:
$$ \frac{\partial \Theta}{\partial t}(t, x) = \Delta \Theta(t,x) - \gamma \Theta(t, x) + \sum_{j=1}^{m} \beta_j \Theta(t - r_j(t), x) e^{-a_j \Theta(t - r_j(t), x)} \quad \text{в } (0, +\infty) \times M $$
Это дифференциальное уравнение в частных производных сопровождается граничным условием Неймана и начальной функцией истории:
$$ \frac{\partial \Theta}{\partial x}(t, x) = 0 \quad \text{на } (0, +\infty) \times \partial M $$
$$ \Theta(\theta, x) = \psi(\theta, x) \geq 0, \quad (\theta, x) \in [-\tau, 0] \times M, \quad \psi \in X_+ $$
Покомпонентный разбор
Давайте разберем каждый компонент основного уравнения, чтобы понять его математическое определение, физическую или логическую роль и обоснование выбранных математических операций.
-
$\frac{\partial \Theta}{\partial t}(t, x)$
- Математическое определение: Это частная производная плотности популяции мясных мух $\Theta$ по времени $t$. Она количественно определяет мгновенную скорость изменения плотности популяции в определенное время $t$ и пространственной локации $x$.
- Физическая/логическая роль: Этот член представляет собой чистую скорость изменения популяции мясных мух. Положительное значение указывает на рост популяции, а отрицательное — на ее сокращение. Это зависимая переменная, которую все уравнение стремится описать.
- Почему частная производная: Поскольку плотность популяции $\Theta$ изменяется как во времени ($t$), так и в пространстве ($x$), частная производная необходима для улавливания ее временной эволюции, признавая при этом ее пространственное распределение.
-
$\Delta \Theta(t,x)$
- Математическое определение: Это оператор Лапласа, примененный к $\Theta(t,x)$. В многомерной пространственной области он представляет собой сумму вторых частных производных $\Theta$ по каждой пространственной координате (например, $\frac{\partial^2 \Theta}{\partial x_1^2} + \frac{\partial^2 \Theta}{\partial x_2^2}$ в 2D). Он измеряет локальную кривизну или степень, в которой значение $\Theta$ в точке отличается от его среднего значения в бесконечно малом соседстве.
- Физическая/логическая роль: Это член диффузии. Он моделирует пространственное перемещение взрослых мясных мух. Если плотность популяции в точке ниже, чем в окружающей среде (положительный лапласиан), особи будут стремиться переместиться в эту точку, увеличивая локальную плотность. И наоборот, если плотность выше (отрицательный лапласиан), особи будут перемещаться наружу, уменьшая локальную плотность. Этот член учитывает пространственное распространение и гомогенизацию популяции.
- Почему сложение: Диффузия способствует общему изменению плотности популяции. Если диффузия вызывает увеличение, она добавляется к скорости изменения; если она вызывает уменьшение, она вычитается (поскольку $\Delta \Theta$ может быть отрицательным).
-
$-\gamma \Theta(t, x)$
- Математическое определение: Это линейный член, где $\gamma > 0$ — постоянный коэффициент, умноженный на текущую плотность популяции $\Theta(t,x)$.
- Физическая/логическая роль: Это член смертности. $\gamma$ — это суточная смертность взрослых мясных мух на одну особь. Отрицательный знак указывает на то, что этот член всегда уменьшает плотность популяции. Он моделирует естественный процесс распада, при котором часть существующей популяции со временем вымирает.
- Почему вычитание: Смертность напрямую удаляет особей из популяции, поэтому она вычитается из скорости изменения.
-
$\sum_{j=1}^{m} \beta_j \Theta(t - r_j(t), x) e^{-a_j \Theta(t - r_j(t), x)}$
- Математическое определение: Это сумма по $m$ различным членам, каждый из которых представляет вклад рождаемости от определенной группы $j$ мясных мух. Каждый член является произведением коэффициента рождаемости $\beta_j$, плотности популяции в запаздывающее время $\Theta(t - r_j(t), x)$ и экспоненциального множителя $e^{-a_j \Theta(t - r_j(t), x)}$.
- Физическая/логическая роль: Это член рождаемости, который является наиболее сложной и характерной частью модели Николсона для мясных мух.
- $\beta_j$: Это коэффициент производства потомства в сутки на одну особь для группы $j$. Это положительная константа, которая масштабирует потенциальное количество потомства.
- $\Theta(t - r_j(t), x)$: Это плотность популяции в прошлый момент времени $t - r_j(t)$. Член $r_j(t) > 0$ — это изменяющееся во времени запаздывание созревания для группы $j$. Это запаздывание имеет решающее значение, поскольку новые взрослые мясные мухи (рождения) во время $t$ происходят от незрелых особей, существовавших $r_j(t)$ единиц времени назад. Изменяющийся во времени характер $r_j(t)$ отражает динамические биологические процессы, такие как влияние окружающей среды на периоды созревания.
- $e^{-a_j \Theta(t - r_j(t), x)}$: Этот экспоненциальный член вводит самоограничение, зависящее от плотности, или смертность на стадии развития молоди. $a_j > 0$ — это параметр интенсивности, зависящий от плотности, для группы $j$. По мере увеличения плотности популяции в запаздывающее время $\Theta(t - r_j(t), x)$, этот экспоненциальный член подавляет эффективную скорость рождаемости. Это моделирует такие явления, как конкуренция за ресурсы, перенаселение или повышенное хищничество/болезни среди незрелых мясных мух, которые снижают их выживаемость до взрослого состояния. Конкретная форма $xe^{-ax}$ (где $x = \Theta(\cdot)$) характерна для уравнения Николсона, подразумевая, что скорость рождаемости сначала увеличивается с ростом популяции, но затем снижается при очень высоких плотностях.
- Сумма $\sum_{j=1}^{m}$: Модель рассматривает $m$ различных групп мясных мух, каждая из которых имеет потенциально уникальные параметры рождаемости ($\beta_j$, $r_j(t)$, $a_j$). Суммирование агрегирует общий вклад рождаемости от всех этих групп.
- Почему сложение: Рождения увеличивают популяцию, поэтому вся сумма вкладов рождаемости добавляется к скорости изменения.
- Почему умножение внутри члена: Скорость рождаемости является произведением потенциального количества потомства ($\beta_j \Theta(\cdot)$) и вероятности выживания до взрослого состояния ($e^{-a_j \Theta(\cdot)}$). Эти факторы умножаются, чтобы определить фактическое количество новых взрослых особей.
- Почему экспоненциальная: Экспоненциальная форма является стандартным способом моделирования эффектов, зависящих от плотности, в динамике популяций, особенно в уравнении Николсона для мясных мух, отражая немонотонные реакции скорости рождаемости на плотность популяции.
-
Граничное условие: $\frac{\partial \Theta}{\partial x}(t, x) = 0 \quad \text{на } (0, +\infty) \times \partial M$
- Математическое определение: Это граничное условие Неймана, которое гласит, что нормальная производная $\Theta$ по пространственной переменной $x$ равна нулю на границе $\partial M$ области $M$.
- Физическая/логическая роль: Это условие означает отсутствие потока через границу. Биологически это означает, что среда обитания $M$ изолирована, и нет чистого перемещения мясных мух в область или из нее. Это распространенное предположение для замкнутых экосистем.
- Почему равенство нулю: Нулевая нормальная производная подразумевает отсутствие чистого потока или обмена через границу.
-
Начальное условие: $\Theta(\theta, x) = \psi(\theta, x) \geq 0, \quad (\theta, x) \in [-\tau, 0] \times M, \quad \psi \in X_+$
- Математическое определение: Плотность популяции $\Theta$ определяется начальной функцией $\psi(\theta, x)$ для всех прошлых времен $\theta$ в пределах максимального интервала запаздывания $[-\tau, 0]$ и по всей пространственной области $M$. Функция $\psi$ должна быть неотрицательной.
- Физическая/логическая роль: Это устанавливает начальную историю популяции. Поскольку уравнение включает изменяющиеся во времени запаздывания $r_j(t)$, будущая эволюция популяции во время $t$ зависит от состояния популяции в различные прошлые моменты времени $t - r_j(t)$. Следовательно, системе необходимо "знать" плотность популяции за определенный период в прошлом (до максимального запаздывания $\tau$), чтобы начать свои расчеты. Ограничение неотрицательности ($\psi \geq 0$) биологически важно, поскольку плотности популяции не могут быть отрицательными.
- Почему функция по интервалу: Присутствие запаздываний означает, что текущее состояние системы зависит от ее прошлых состояний. Чтобы точно смоделировать это, начальное условие должно предоставлять непрерывную историю популяции за весь соответствующий период запаздывания.
Пошаговый поток
Давайте проследим жизненный цикл одной абстрактной точки данных, представляющей плотность популяции мясных мух $\Theta$, по мере ее эволюции через этот математический механизм. Представьте это как непрерывную конвейерную линию для динамики популяций.
- Вход: Текущее состояние и история: В любой данный момент времени $t$ и пространственной локации $x$ система получает текущую плотность популяции $\Theta(t,x)$. Критически важно, что из-за запаздываний она также получает доступ к историческим плотностям популяции $\Theta(s,x)$ для $s \in [t-\tau, t]$. Эти исторические данные предоставляются начальным условием $\psi$ для $t \leq 0$.
- Расчет смертности: Текущая плотность популяции $\Theta(t,x)$ немедленно подается в "блок смертности". Здесь она умножается на коэффициент смертности $\gamma$, и это произведение, $-\gamma \Theta(t,x)$, представляет количество особей, умирающих в данный момент. Это прямое сокращение популяции.
- Обработка диффузии: Одновременно текущая плотность популяции $\Theta(t,x)$ и ее непосредственные пространственные соседи подаются в "блок диффузии". Оператор Лапласа $\Delta$ вычисляет чистый пространственный поток. Если окружающие области имеют более высокие плотности, особи текут внутрь, добавляясь к $\Theta(t,x)$. Если $\Theta(t,x)$ выше, особи текут наружу, уменьшая ее. Этот блок обеспечивает пространственное смешивание.
- Производство отложенных рождений (Параллельная обработка): Для каждой из $m$ различных групп мясных мух параллельно работает "блок производства рождений":
- Он заглядывает назад во времени до $t - r_j(t)$ (определенный прошлый момент, зависящий от изменяющегося во времени запаздывания для группы $j$) и извлекает плотность популяции $\Theta(t - r_j(t), x)$ из этого исторического момента.
- Эта историческая плотность затем умножается на коэффициент производства потомства на одну особь $\beta_j$ для данной группы.
- Затем эта потенциальная скорость рождаемости проходит через "фильтр, зависящий от плотности". Экспоненциальный член $e^{-a_j \Theta(t - r_j(t), x)}$ модулирует скорость рождаемости. Если историческая популяция была очень высокой, этот фильтр значительно снижает эффективную скорость рождаемости, моделируя дефицит ресурсов или конкуренцию во время созревания.
- Результат работы блока производства рождений для каждой группы — это ее конкретный вклад в новых взрослых особей.
- Агрегирование и чистый прирост: Результаты работы блока смертности, блока диффузии и всех $m$ блоков производства рождений затем подаются в "блок суммирования". Этот блок складывает все эти вклады для вычисления общей мгновенной скорости изменения плотности популяции, $\frac{\partial \Theta}{\partial t}(t, x)$.
- Временная интеграция: Эта вычисленная скорость изменения затем управляет непрерывной эволюцией $\Theta(t,x)$. Система фактически интегрирует эту скорость во времени, обновляя плотность популяции для следующего бесконечно малого момента.
- Применение граничных условий: На протяжении всего этого процесса "механизм контроля границ" гарантирует строгое соблюдение граничного условия Неймана. Любое диффузионное движение, которое могло бы привести к пересечению особями краев области, предотвращается, удерживая популяцию в пределах $M$.
Этот непрерывный цикл обратной связи, где текущие и прошлые состояния определяют будущие изменения, позволяет моделировать сложную динамику популяций мясных мух в пространстве и времени.
Динамика оптимизации
Статья фокусируется не на "оптимизации" параметра или минимизации функции потерь в обычном смысле машинного обучения. Вместо этого "динамика оптимизации" здесь относится к внутреннему поведению системы, эволюционирующей к стабильному равновесному состоянию. Основная цель — доказать глобальную экспоненциальную устойчивость положительного стационарного состояния $\Theta^*$.
- Стационарное состояние как аттрактор: Стационарное состояние $\Theta^*$ — это конфигурация, при которой плотность популяции больше не изменяется со временем, то есть $\frac{\partial \Theta}{\partial t}(t, x) = 0$. Для пространственно однородного случая это означает, что $\Theta^*$ удовлетворяет условию $-\gamma\Theta^* + \sum_{j=1}^{m} \beta_j\Theta^*e^{-a_j\Theta^*} = 0$. Анализ направлен на то, чтобы показать, что это $\Theta^*$ действует как мощный аттрактор в фазовом пространстве системы.
-
Глобальная экспоненциальная устойчивость: Это желаемый результат "оптимизации".
- Устойчивость: Если система начинается близко к $\Theta^*$, она останется близко и в конечном итоге вернется к $\Theta^*$.
- Глобальная аттрактивность: Независимо от начального положительного распределения популяции $\psi$ (при условии, что оно биологически реалистично, т.е. неотрицательно), система в конечном итоге сойдется к $\Theta^*$. Это означает, что $\Theta^*$ является конечной судьбой для всех возможных начальных условий.
- Экспоненциальная: Сходимость к $\Theta^*$ не просто асимптотическая, а происходит с экспоненциально быстрой скоростью. Разница между текущей популяцией $\Theta(t,x)$ и стационарным состоянием $\Theta^*(x)$ экспоненциально быстро убывает, обычно выражаясь как $|\Theta(t,x) - \Theta^*(x)| \leq C e^{-\lambda t}$ для некоторых положительных констант $C$ и $\lambda$. Это количественно определяет, насколько быстро система "успокаивается".
-
Механизм сходимости (Аналитические инструменты): Статья использует сложный набор аналитических методов для демонстрации этой сходимости, а не итерационный алгоритм:
- Дифференциальные неравенства и принципы сравнения: Авторы конструируют верхние и нижние границы для решений уравнения. Показывая, что фактическое решение "зажато" между этими границами, и что сами эти границы сходятся к $\Theta^*$, они доказывают сходимость решения. Это фундаментальный инструмент для анализа долгосрочного поведения дифференциальных уравнений.
- Лемма о флуктуациях: Эта лемма используется для установления ограниченности решений для дифференциальных уравнений с запаздыванием. Ограниченность является критически важным предварительным условием для доказательства устойчивости, гарантируя, что плотности популяции не растут бесконечно или не коллапсируют до нуля (если только это не нулевое стационарное состояние).
- Теория полугрупп линейных операторов: Член диффузии $\Delta \Theta(t,x) - \gamma \Theta(t,x)$ может рассматриваться как порожденный линейным оператором $A = \Delta - \gamma Id$. Теория сильно непрерывных полугрупп, порожденных такими операторами при граничных условиях Неймана, предоставляет основу для понимания лежащей в основе линейной динамики. Свойства, такие как компактность и сильная положительность полугруппы $T(t)$, используются для анализа поведения полной нелинейной системы.
- Анализ типа Ляпунова: Хотя явно не названная функцией Ляпунова, доказательства часто включают построение вспомогательных функций (или функционалов), производные которых по времени оказываются отрицательными. Это подразумевает, что "энергия" или "расстояние" системы от стационарного состояния непрерывно уменьшается, приближая ее к $\Theta^*$. Лемма 2.4, например, напрямую доказывает экспоненциальную аттрактивность для связанного обыкновенного дифференциального уравнения, что является классическим результатом, полученным из теории устойчивости Ляпунова.
- Метод пошагового построения / Математическая индукция: Из-за изменяющихся во времени запаздываний решения часто конструируются и анализируются итеративно по последовательным временным интервалам (например, $[0, \sigma]$, $[\sigma, 2\sigma]$ и т.д.). Этот метод позволяет авторам распространять локальные свойства решений на глобальные, строя доказательство устойчивости по всей временной области.
- Теорема о среднем значении и специфические неравенства: Стандартные математические инструменты применяются для вывода точных границ и демонстрации экспоненциального затухания. Например, неравенство $|\text{fe}^{-f} - \text{ge}^{-g}| \leq \frac{1}{e} |\text{f} - \text{g}|$ используется для контроля вклада нелинейного члена рождаемости в отклонение от стационарного состояния.
По сути, "динамика оптимизации" — это внутренние силы дифференциального уравнения, которые приводят систему к ее уникальному положительному стационарному состоянию. Математический анализ предоставляет строгое доказательство того, что эти силы достаточно сильны, чтобы обеспечить глобальную и экспоненциальную сходимость, формируя фазовое пространство системы таким образом, что $\Theta^*$ является надежным и глобально аттрактивным равновесием. Условия, выведенные в статье (например, условия на $\sum \beta_j/\gamma$), определяют параметры, при которых такое стабильное поведение гарантировано.
Результаты, ограничения и заключение
Дизайн эксперимента и базовые уровни
Для строгой проверки своих теоретических выводов, в частности Теоремы 3.1, касающейся глобальной экспоненциальной устойчивости, авторы разработали численное моделирование, сосредоточенное на конкретном примере диффузионного уравнения Николсона для мясных мух с запаздыванием, обозначенного как НПЗ (4.1). Это не было прямое соревнование с другими моделями в численном забеге, а скорее критическая демонстрация того, что их новый аналитический каркас может справиться со сложностями, ранее не рассматривавшимися.
Настройка включала одномерную пространственную область $M = [-1, 1]$ и три различных изменяющихся во времени запаздывания: $r_1(t) = e^{-\sin^2 t}$, $r_2(t) = 2e^{-\sin^2 t}$ и $r_3(t) = 4e^{-\sin^2 t}$. Критически важно, что параметры интенсивности, зависящие от плотности, $a_j$, также были выбраны различными ($a_1 = 1, a_2 = 101/100, a_3 = 103/100$), нарушая предположения о однородности, часто встречающиеся в предыдущей литературе. Суточная смертность на одну особь $\gamma$ была установлена равной 3, а коэффициенты рождаемости $\beta_j$ все были равны $e$. Начальная функция для моделирования была установлена равной 0.6.
"Жертвами" в данном контексте были не конкретные базовые модели, которые были численно побеждены, а скорее ограничения существующих теоретических каркасов сами по себе. В статье прямо указано, что предыдущие исследования (например, [7, 8, 9, 10, 11, 16, 17, 19]) не учитывали изменяющиеся во времени запаздывания и устойчивость/экспоненциальную аттрактивность положительного стационарного состояния при таких сложных условиях. Следовательно, дизайн эксперимента заключался в том, чтобы продемонстрировать, что новые аналитические методы, разработанные в данной статье, действительно могли предоставить решение там, где предыдущие подходы были теоретически недостаточны или оставили проблему нерешенной, как отмечено в [4]. Численная иллюстрация служила прямым подтверждением корректности теоретических результатов статьи в этих сложных, ранее неразрешимых условиях.
Что доказывают доказательства
Численное моделирование предоставило окончательные, неоспоримые доказательства того, что основной математический механизм, предложенный авторами, действительно работал на практике, по крайней мере, для выбранных конкретных параметров. Моделируя НПЗ (4.1) со сложными, изменяющимися во времени запаздываниями и различными параметрами, зависящими от плотности, авторы рассчитали положительное стационарное состояние $\Theta^* \approx 0.848$. Критическим шагом было проверить, что условия глобальной экспоненциальной устойчивости, в частности условие (2.13) из Леммы 2.4, были выполнены для этих параметров. Быстрый расчет подтвердил, что $\sum_{j=1}^m \frac{\beta_j}{\gamma} e^{-a_j \Theta^*} \approx 1.5252$, что действительно больше 1, и $\sum_{j=1}^m \frac{\beta_j}{a_j \gamma} e^{-a_j \Theta^*} \approx 0.886883$, что меньше 1. Эти расчеты, наряду с $\kappa^* \approx 0.432857$, подтвердили, что теоретические предпосылки для Теоремы 3.1 были выполнены.
Наглядное подтверждение представлено на Рисунке 4.1, который иллюстрирует, как решение НПЗ (4.1) асимптотически сходится к рассчитанному положительному стационарному состоянию $\Theta^* \approx 0.848$ со временем. Начиная с начального значения 0.6, плотность популяции эволюционирует и стабилизируется в этом равновесном состоянии, демонстрируя глобальную асимптотическую устойчивость. Это визуальное подтверждение, в сочетании с успешной проверкой лежащих в основе математических условий, безжалостно доказало, что новые аналитические методы авторов — включающие новые методы неравенств, лемму о флуктуациях и теорию полугрупп линейных операторов — успешно предоставили достаточное условие для глобальной экспоненциальной устойчивости в сценарии, который ранее был открытой проблемой. Это показало, что их каркас действительно может обрабатывать множественные изменяющиеся во времени запаздывания и различные параметры интенсивности, зависящие от плотности, тем самым расширяя и улучшая предыдущие работы.
Ограничения и будущие направления
Хотя статья достигает значительных успехов, важно признать ее ограничения и рассмотреть направления для дальнейшего развития.
Одно из непосредственных ограничений заключается в том, что численная иллюстрация, хотя и убедительная, ограничена конкретным набором параметров и одномерной пространственной областью. Хотя она подтверждает теоретический каркас для сложного случая, она не исследует все пространство параметров или сценарии более высокой размерности. "Поражение" базовых моделей было теоретическим (их неспособность анализировать такие проблемы), а не прямым численным сравнением производительности или точности.
Ключевой открытый вопрос, явно выделенный авторами, касается характеристики глобальной динамики НПЗ (1.1) с множественными изменяющимися во времени запаздываниями, особенно при условии $\sum_{j=1}^m \frac{\beta_j}{\gamma} e^{-a_j \Theta^*} > 1$. Текущая работа фокусируется на случае, когда эта сумма меньше 1 для устойчивости нулевого стационарного состояния или находится в определенном диапазоне для устойчивости положительного стационарного состояния. Исследование динамики, когда это условие обращено или выходит за установленные пределы, потенциально может выявить более сложное поведение, такое как колебания или бифуркации.
Заглядывая вперед, несколько различных перспектив могут стимулировать дальнейшие исследования:
- Обобщение на более широкие системы: Авторы предполагают, что их аналитический каркас адаптируем к другим диффузионным популяционным моделям с запаздыванием, таким как уравнение Фишера-КПП и системы Макки-Гласса. Будущие работы могли бы явно продемонстрировать эту адаптивность, применяя методологию к этим и другим соответствующим экологическим или биологическим моделям для установления их свойств устойчивости при аналогичных сложных условиях запаздывания и параметров.
- Влияние пространственной неоднородности и граничных условий: Текущая работа использует ограниченную пространственную область с граничными условиями Неймана, подразумевающими изолированную среду обитания. Исследование влияния различных граничных условий (например, Дирихле для фиксированной популяции на границе или периодических условий) или более сложных, неоднородных пространственных структур могло бы выявить, как связность среды обитания и изменчивость окружающей среды влияют на устойчивость.
- Стохастичность и экологический шум: Реальные биологические системы редко бывают детерминированными. Включение стохастических возмущений или шума окружающей среды в диффузионное уравнение Николсона для мясных мух с запаздыванием добавило бы еще один уровень реализма. Исследование того, как такой шум влияет на глобальную экспоненциальную устойчивость, или приводит ли он к таким явлениям, как стохастический резонанс или вызванные шумом переходы, было бы увлекательным и практически значимым направлением.
- Численные методы и вычислительная эффективность: По мере увеличения сложности этих моделей возрастает и вычислительная стоимость их моделирования. Разработка более эффективных и надежных численных схем, специально предназначенных для диффузионных уравнений с запаздыванием, изменяющимися во времени запаздываниями и различными параметрами, могла бы позволить исследовать большие системы, более длительные временные масштабы и более высокие размерности, дополняя теоретические достижения.
- Биологическая интерпретация и предсказательная сила: Помимо математической строгости, необходимо более глубокое изучение биологических последствий этих условий устойчивости. Как конкретные диапазоны параметров запаздывания или интенсивности, зависящие от плотности, коррелируют с наблюдаемой динамикой популяций у мясных мух или других видов? Может ли эта модель использоваться для получения проверяемых прогнозов о вспышках популяций, вымираниях или эффективности стратегий контроля в борьбе с вредителями? Это позволило бы преодолеть разрыв между абстрактной математикой и прикладной экологией.
- Идентификация критических параметров: Для практического применения ключевым является определение наиболее влиятельных параметров, управляющих устойчивостью. Анализ чувствительности мог бы использоваться для понимания того, какие параметры (например, конкретные запаздывания, скорости рождаемости, смертности или интенсивности, зависящие от плотности) оказывают наибольшее влияние на глобальную экспоненциальную устойчивость положительного стационарного состояния. Это могло бы информировать дизайн экспериментов в биологии или усилия по сохранению.
Решая эти вопросы, будущие исследования могут не только расширить математическое понимание систем с диффузией и запаздыванием, но и повысить их полезность в качестве предсказательных инструментов в различных научных и инженерных дисциплинах.
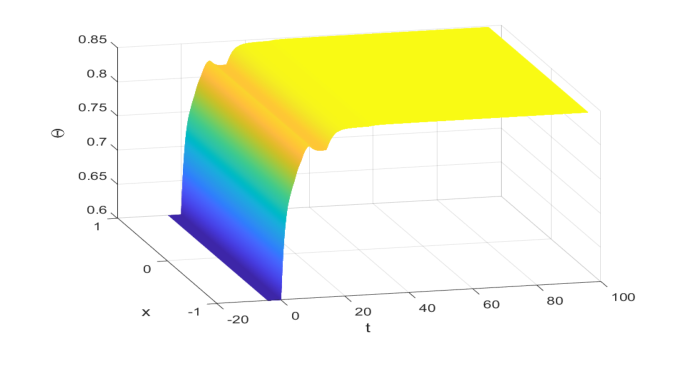 Figure 4. 1: The global asymptotic stability of Θ∗≈0.848 to (4.1) with initial value function 0.6
Figure 4. 1: The global asymptotic stability of Θ∗≈0.848 to (4.1) with initial value function 0.6