二重飽和変換項を有する植生水モデルの動的挙動解析
In this paper, we propose a vegetation-water system incorporating double saturation transformation terms, which more vividly depicts the mutual influence and transformation relationship between vegetation and water.
背景と学術的系譜
起源と学術的系譜
本研究の正確な起源は、乾燥、半乾燥、半湿潤地域に広く存在する重要な環境問題である土地砂漠化の問題にあります。これは、自然要因と人為活動によって砂漠化が加速され、土壌浸食、植生減少、生態系安定性の低下を引き起こす乾燥、半乾燥、半湿潤地域で世界的に発生しています。数理生態学の分野は、これらの複雑な相互作用をモデル化しようと試みてきました。
歴史的には、Klausmeier [8] が空間拡散と輸送を組み込んだ基本的な植生-水システム(モデル1.1)を導入しました。このモデルは、その後の研究の基礎となりました。例えば、Sherrattら [9] は断続的な降雨を研究するためにインパルスフレームワークを統合し、Consoloら [10] は一般化された双曲線型Klausmeierモデルを用いてチューリングパターンを解析しました。Kealyら [11] は、対流項を除去し線形拡散を用いることでKlausmeierモデル(モデル1.2につながる)を単純化し、平坦な地形への適用性を高めました。この単純化されたモデルは、Sunら [12]、Zhangら [13]、Gouら [14] によって、安定な配置、分岐パラメータ、植生分布へのパラメータの影響を特徴づけるためにさらに調査されました。その後、Liら [29] は、植物が生理学的に水の吸収を調節し、特定のバイオマス閾値で最大吸収能力に達することを認識して、水吸収飽和効果をシステム(モデル1.3)に導入しました。
モデル(1.3)を含むこれらの先行アプローチの根本的な限界、すなわち「ペインポイント」は、植生と水の間の双方向飽和相互作用を完全に捉えられないことでした。モデル(1.3)は植生の飽和水吸収を考慮しましたが、水利用可能性自体の飽和特性を明示的にモデル化していませんでした。この省略は、以前のモデルが、特に水と植生の両方が同時に制限要因となりうる砂漠化しやすい地域において、植生と水の相互影響と変換関係の、より鮮明で生物学的に忠実な描写を提供できないことを意味しました。このギャップにより、著者らは、この重要な双方向フィードバックに対処するために「二重飽和変換項」を組み込んだ現在の論文のモデルを開発せざるを得なくなりました。これにより、解析の複雑さが増すにもかかわらず、モデルの生物学的忠実性が向上しました。
直感的な領域用語
- 土地砂漠化: かつて緑豊かだった庭が、ゆっくりと不毛の砂漠地帯に変わっていく様子を想像してください。これは、自然の乾燥と過放牧や森林破壊のような人為活動の組み合わせにより、土壌が植物の生育を支える能力を失ったときに起こります。まるで庭がゆっくりと「乾いて」砂漠になっているかのようです。
- チューリング不安定性: 均一な色のケーキ生地を想像してください。特定の材料(反応における特定の化学物質や生物学的システムにおける拡散率など)を導入し、それらを相互作用させると、生地が固まるにつれて、ストライプ、斑点、迷路のような、明確で繰り返されるパターンが自発的に発生する可能性があります。チューリング不安定性は、均一な状態が不安定になり、複雑で組織化されたパターンを生み出すこの現象を説明します。
- 二重飽和変換項: 喉が渇いた植物(植生)が水源から水を飲もうとしていると考えてください。まず、植物自体は、どれだけ水があっても吸収できる量に限りがあります(植生飽和)。次に、土壌に元々水が非常に少ない場合、植物はより多く吸収できる能力があっても、あまり吸収できません(水飽和)。モデルにおけるこの「二重飽和」項は、これら両方の制限を同時に考慮し、相互作用をより現実的にしています。
- 分岐現象: 一本の安定した流れで流れる川を想像してください。土地の傾斜や流れる水の量のような条件を徐々に変えると、川は突然二つの異なる水路に分かれたり、劇的に蛇行し始めたり、あるいは干上がったりするかもしれません。数学では、分岐とは、システムパラメータの小さな連続的な変化が、その挙動またはその解の数と種類に突然の質的な変化をもたらすことです。
記法表
| 記法 | 説明 | タイプ |
|---|---|---|
| $w$ | 無次元化された水の密度 | 変数 |
| $n$ | 無次元化された植生バイオマス | 変数 |
| $x$ | 無次元化された空間座標 | 変数 |
| $t$ | 無次元化された時間 | 変数 |
| $s$ | 無次元化された水の収量(降雨強度) | パラメータ |
| $p$ | 植生成長/水変換に関連する無次元化されたパラメータ | パラメータ |
| $b$ | 無次元化された植生減少率 | パラメータ |
| $k$ | 無次元化された水の飽和閾値 | パラメータ |
| $d_1$ | 無次元化された土壌水拡散係数 | パラメータ |
| $d_2$ | 無次元化された植生拡散係数 | パラメータ |
問題定義と制約
中核問題の定式化とジレンマ
本論文が取り組む中核的な問題は、砂漠化しやすい地域における植生-水相互作用のための、より生物学的に忠実な数学的モデルを開発することです。Klausmeierの(1.1)や、Kealyら(1.2)やLiら(1.3)のようなその拡張のような以前のモデルは、植生の水の吸収能力の飽和効果を捉えていましたが、それは $F(N) = \frac{aN^2}{1+bN}$ のような項で表されていました。
入力/現在の状態は、既存の植生-水モデルのセットですが、これらは基礎的であるものの、植生と水との間の双方向飽和相互作用を完全に捉えていません。具体的には、水利用可能性自体の飽和を考慮するメカニズムが欠如しており、生態水文学的フィードバックの現実的でない描写につながっています。
出力/目標状態は、二重飽和変換項を組み込んだ新しい植生-水システム、特にモデル(1.4)とその無次元化された形式(1.5)です。この新しいモデルは、植生と水の間の相互影響と変換関係をより正確かつ鮮明に記述することを目的としており、それによって砂漠化しやすい地域における水分-植生相互作用を表現する上での生物学的忠実性を向上させます。最終的な目標は、これらの洗練された条件下での、大域的安定性、チューリング不安定性、および様々な分岐現象を含む動的挙動の、より深く現実的な解析を可能にすることです。
本論文が橋渡ししようとしている正確に欠けているリンクまたは数学的なギャップは、水の飽和ダイナミクスにあります。以前のモデルは植生の水の吸収の飽和($F(N)$)を考慮していましたが、水自体の飽和を明示的にモデル化していませんでした。本論文は、水飽和関数 $G(W) = \frac{W}{W+K}$ (ここで $W$ は水の密度、$K$ は飽和定数)を導入します。重要な革新は、複合的な二重飽和項 $RG(W) \cdot F(N) \cdot N$ (またはその無次元化された等価物 $\frac{pwn^2}{(w+k)(n+1)}$)です。この項は、植生の飽和水吸収能力と水自体の飽和特性の相乗的な制限を正確に特徴づけ、非線形生態水文学的フィードバックのより正確な解明を提供します。
以前の研究者を閉じ込めてきた痛みを伴うトレードオフまたはジレンマであり、著者らによって明示的に強調されているのは、生物学的現実性と解析的実行可能性との間のバランスです。「水飽和関数G(W)の包含は、モデルの生物学的忠実性を大幅に向上させ、砂漠化しやすい地域における水分-植生相互作用をより現実的に記述することを可能にします。しかし、高次の非線形性の導入により、定常状態解の導出と分岐解析の実施において、解析の複雑さも増大します。」これは、より正確な生物学的表現を達成することが、システムを解き解析する上での数学的困難の増大という重大なコストを伴うことを意味します。
制約と失敗モード
砂漠化しやすい地域における植生-水ダイナミクスを正確にモデル化するという問題は、著者らが直面するいくつかの厳しい現実的な壁のために、非常に困難です。
-
物理的および生態学的制約:
- 非負解: どの生物学的モデルにおいても、水($w$)と植生($n$)の密度は非負でなければなりません。システム(1.5)は、非負の初期データ $(w_0(x,0), n_0(x,0)) \ge (0,0)$ の制約を受けますが、これは物理的現実性の基本的な要件です。非負性を維持できないと、モデルは生物学的に意味をなさなくなります。
- 有界領域とゼロフラックス境界条件: 解析は、滑らかな境界を持つ有界空間領域 $\Omega$ 上で、ゼロフラックス(ノイマン)境界条件($\frac{\partial w}{\partial \nu} = \frac{\partial n}{\partial \nu} = 0$ on $\partial \Omega$)の下で行われます。これらの条件は、水や植生が境界を通過しない閉鎖システムを表しており、多くの生態学的パッチにとって現実的な仮定ですが、偏微分方程式(PDE)に複雑さを加えます。
- 飽和現象: 問題の核心は、植生の水の吸収と水利用可能性自体の両方の飽和を正確に捉えることです。これは、モデルに組み込まれなければならない複雑な生理学的および環境的現実です。
- 環境要因: モデルは、植生の分布と水のダイナミクスに影響を与える様々な自然要因(降雨、風速、光)と人為活動(畜産)を暗黙的または明示的に考慮する必要があります。
-
計算上および数学上の制約:
- 高次の非線形性: 二重飽和項 $G(W) \cdot F(N)$ の導入は、システム(1.4)とその無次元化された形式(1.5)の非線形性を大幅に増加させます。これにより、線形または低次の非線形システムと比較して、定常状態解の導出、安定性解析、および分岐解析が著しく困難になります。標準的な解析手法は、このような高次の非線形性にはしばしば苦労します。
- 定常状態解の解析的複雑さ: 高度に非線形な反応拡散系の明示的な定常状態解を見つけることは、一般的に非常に困難です。本論文は、厳密な数学的導出、事前推定、および定性分析に依存していますが、これらは計算集約的であり、高度な数学的ツールを必要とします。
- 分岐解析の課題: 単純および二重固有値に関連する分岐現象の解析には、Lyapunov-Schmidt還元法やCrandall-Rabinowitz定理のような洗練された方法が必要です。これらの方法は複雑であり、非線形性が増すとその適用はより複雑になります。本論文では、特定の条件(例えば、$j \neq 2i$ かつ $i \neq 2j$ の場合)では、陰関数定理を適用して解の存在を確立できないと指摘しており、標準的な解析アプローチの失敗モードを示唆しています。
- チューリング不安定性の条件: チューリング不安定性(パターン形成につながる)が発生する条件を決定するには、線形化されたシステムの複雑な固有値解析が必要であり、これは拡散係数やその他のパラメータに強く依存します。
- パラメータ空間の探索: モデルには多数のパラメータ(例:$s, p, b, k, d_1, d_2$)が含まれています。各パラメータがシステムの動的挙動とパターン形成に与える影響を徹底的に調査するには、広範な理論的解析と数値シミュレーションが必要であり、問題空間が広大で完全に探索するのが困難になっています。本論文の数値シミュレーションは、この複雑なパラメータ空間全体にわたる理論的予測を検証するために不可欠です。
なぜこのアプローチなのか
選択の必然性
本論文の著者らは、既存の植生-水モデルに根本的な限界があることに直面しました。それは、砂漠化しやすい地域における植生と水の間の複雑な双方向飽和効果を十分に捉えられないことでした。Klausmeierのシステム(システム1.1)や、Kealyらのシステム(システム1.2)、Liらのシステム(システム1.3)のような従来のモデルは、主に単一の飽和項を組み込んでおり、通常はバイオマスが増加するにつれて植生の水の吸収能力が飽和する様子を表していました。これは、$f(N) = \frac{aN^2}{1+bN}$ のような関数で表されます。
しかし、著者らはこれが不十分であることに気づきました。この認識の正確な瞬間は、3ページで次のように述べられています。「砂漠化しやすい地域における植生と水の間の双方向飽和相互作用をより良く捉えるために、二重飽和変換メカニズムを組み込むことによってシステム(1.3)をさらに洗練させます。」これは、問題が単に植生が水を吸収することだけでなく、水自体の飽和特性と、それが同時に植生成長をどのように制限するかということでもあることを強調しています。両方を考慮しなければ、モデルは生態学的なパズルの重要なピースを欠いていました。したがって、二重飽和項を組み込んだモデルは、単なる改善ではなく、これらの複雑な相互制限を正確に表現するための唯一実行可能な解決策でした。これは明確な短所でした。
比較優位性
このアプローチの質的な優位性は、単に計算上のパフォーマンス指標の改善ではなく、生物学的忠実性の向上に直接由来します。新しい非線形連鎖項 $RG(W) \cdot F(N) \cdot N$ を介した「二重飽和変換項」の導入が、主要な構造的利点です。この項は、3ページで説明されているように、「吸収ダイナミクスにおける水と植生の同時制限を特徴づける」ものです。
単一飽和のみを考慮した以前のモデルは、この相乗的な制限を完全に表現できませんでした。植生吸収率 $F(N)$ とともに水飽和関数 $G(W)$ を含めることにより、モデルは「砂漠化しやすい地域における水分-植生相互作用をより現実的に記述する」ことができ、「非線形生態水文学的フィードバックのメカニズムをより正確に解明する」ことができます。これは、モデルが単にデータをより良く適合させているのではなく、現実世界の生態学的プロセスをより忠実に反映するように構造化されていることを意味します。それは、これらの脆弱な環境でパターン形成と生態系安定性を駆動する複雑なダイナミクスを、先行するゴールドスタンダードよりも大幅な質的飛躍となる、より深く、より微妙な理解を提供します。本論文は、その焦点がメカニズム的な微分方程式モデリングアプローチにあるため、高次元ノイズの処理やメモリ複雑性の削減のような側面については議論していません。
制約との整合性
正直なところ、ステップ2で定義された特定の制約については、そのセクションが提供されなかったため、完全には確信が持てません。しかし、論文で提示された問題定義と動機に基づいて、この選択された方法が完全に満たすいくつかの厳しい要件を推測できます。中核的な問題は、「砂漠化しやすい地域」における植生-水相互作用の正確なモデリングを中心に展開しており(要旨、3ページ)、これは環境ストレス下での重要なフィードバックメカニズムを捉える上での高い忠実性の必要性を示唆しています。
問題の要件と解決策のユニークな特性との「結婚」は、いくつかの点で明らかです。
1. 相互影響の現実的な描写: 問題は、「植生と水の間の相互影響と変換関係をより鮮明に描写する」モデルを要求しています(要旨)。二重飽和項、$G(W)$ と $F(N)$ は、水利用可能性と植生の吸収能力の両方が同時に互いを制限する方法を明示的にモデル化することにより、これを直接満たします。
2. 双方向飽和相互作用: 著者らは、「砂漠化しやすい地域における植生と水の間の双方向飽和相互作用をより良く捉える」ことを明確に求めていました(3ページ)。新しい二重飽和項 $RG(W) \cdot F(N) \cdot N$ はまさにこの目的のために設計されており、モデルが相互作用の両側からの飽和を考慮することを保証します。これは、以前のモデルでは重要な欠落部分でした。
3. 非線形メカニズムの解明: 砂漠化は複雑な非線形生態水文学的フィードバックを伴います。選択された方法論は、二重飽和項によって導入される高次の非線形性により、「砂漠化しやすい地域における非線形生態水文学的フィードバックのメカニズムをより正確に解明する」ことを可能にします(3ページ)。これは、これらの脆弱な環境におけるパターン形成と生態系安定性を駆動する複雑なダイナミクスを理解する必要性と完全に一致しています。
代替案の却下
本論文が代替アプローチを却下する理由は、主に暗黙的であり、GANやTransformerのような全く異なる方法論的パラダイムというよりも、以前の数学的モデルの限界に焦点を当てています。著者らは、Klausmeierのシステム(1.1)とその後の拡張(例:Kealyらのシステム(1.2)、Liらのシステム(1.3))から始まる反応拡散モデルの系譜を築いています。
これらの以前のモデルが不十分であると却下する主な理由は、「植生と水の間の双方向飽和相互作用」を捉えられないことでした(3ページ)。これらのモデルは単一の飽和項(例:植生の水の吸収のための $f(N) = \frac{aN^2}{1+bN}$)を組み込んでいましたが、水飽和のための対応する項 $G(W)$ が欠如していました。著者らは、「伝統的な」方法、この文脈では単一飽和植生-水モデルを意味するものが、両方の飽和効果を「同時に捉えることができない」ため、「この特定の問題には不十分」であると見出しました(4ページ)。この省略は、生態水文学的フィードバックの現実的でない描写につながりました。したがって、二重飽和項を持つ新しいモデルは、同じモデリングフレームワーク内での先行モデルのこれらの特定の欠点を克服するために必要な洗練として導入されました。本論文は、GANや拡散モデルのような機械学習モデルが失敗する理由については、それらが異なる抽象化レベルで動作し、ここで行われているメカニズム的な微分方程式モデリングアプローチと直接比較できないため、詳しく説明していません。
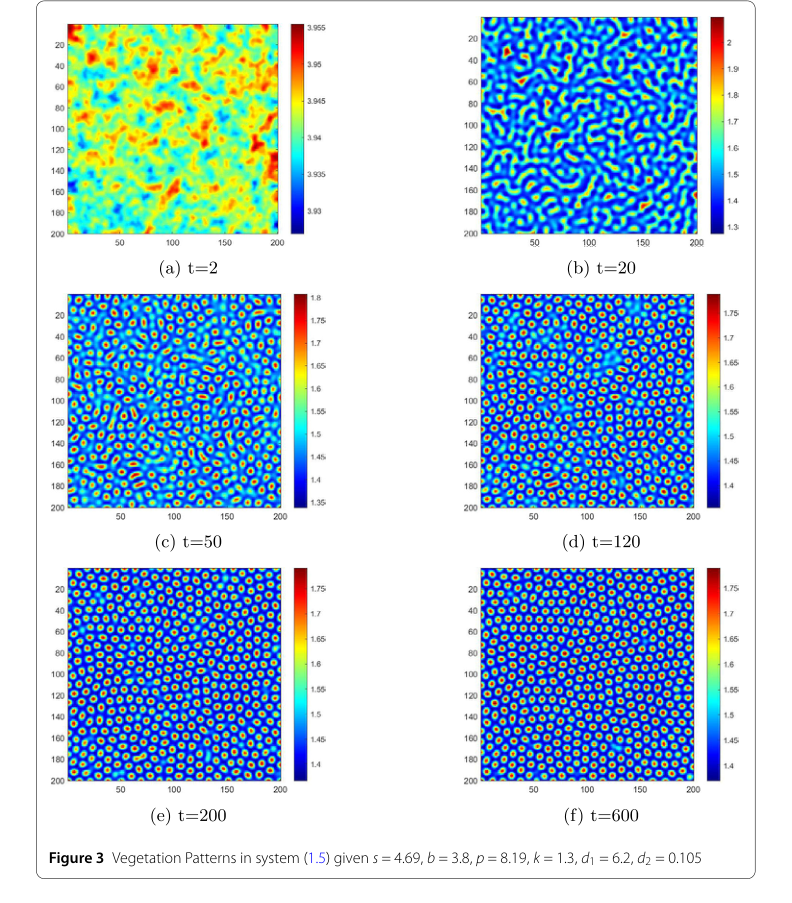 Figure 3. Vegetation Patterns in system (1.5) given s = 4.69, b = 3.8, p = 8.19, k = 1.3, d1 = 6.2, d2 = 0.105
Figure 3. Vegetation Patterns in system (1.5) given s = 4.69, b = 3.8, p = 8.19, k = 1.3, d1 = 6.2, d2 = 0.105
数学的・論理的メカニズム
マスター方程式
本論文の数学的エンジンの絶対的な核心は、植生バイオマスと水の密度の動的な相互作用を記述する、無次元化された反応拡散システムです。このシステムは、二重飽和変換項を組み込んだ元のモデルから導出され、その後スケール化されたもので、次のように提示されます。
$$ \begin{cases} \frac{\partial w}{\partial t} = s - w - \frac{pwn^2}{(w+k)(n+1)} + d_1\Delta w, & x \in \Omega, t > 0, \\ \frac{\partial n}{\partial t} = \frac{pwn^2}{(w+k)(n+1)} - bn + d_2\Delta n, & x \in \Omega, t > 0, \\ \frac{\partial w}{\partial \nu} = \frac{\partial n}{\partial \nu} = 0, & x \in \partial\Omega, t > 0, \\ (w_0(x, 0), n_0(x, 0)) \geq, \neq (0,0), & x \in \Omega. \end{cases} $$
項ごとの解剖
このマスター方程式を一つずつ分解して、その構成要素と役割を理解しましょう。
-
$w$:
- 数学的定義: 無次元化された水の密度を表す従属変数。これはスカラー場であり、その値は空間と時間によって変化する可能性があります。
- 物理的/論理的役割: これは、任意の場所 $x$ と時間 $t$ における利用可能な水の量を定量化します。
- 使用理由: 水の状態変数です。
-
$n$:
- 数学的定義: 無次元化された植生バイオマス密度を表す従属変数。$w$ と同様に、スカラー場です。
- 物理的/論理的役割: これは、任意の場所 $x$ と時間 $t$ における植生バイオマスの量を定量化します。
- 使用理由: 植生の状態変数です。
-
$t$:
- 数学的定義: 時間を表す独立変数。
- 物理的/論理的役割: これは、システムが進化する時間的次元です。
- 使用理由: $w$ と $n$ が時間とともにどのように変化するかを記述するため。
-
$x \in \Omega, t > 0$:
- 数学的定義: 偏微分方程式(PDE)の領域を指定します。$\Omega$ は有界な空間領域であり、$t > 0$ は初期時点以降のシステムの進化を見ていることを意味します。
- 物理的/論理的役割: モデルが適用される物理空間と時間間隔を定義します。
- 使用理由: 空間的および時間的ダイナミクスの文脈を設定するため。
-
$\frac{\partial w}{\partial t}$:
- 数学的定義: 時間 $t$ に関する水の密度 $w$ の偏微分。
- 物理的/論理的役割: これは、空間の特定点における水の密度の瞬間的な変化率を表します。正の値は水の増加を意味し、負の値は減少を意味します。
- 使用理由: 水の時間的進化をモデル化するため。
-
$\frac{\partial n}{\partial t}$:
- 数学的定義: 時間 $t$ に関する植生バイオマス密度 $n$ の偏微分。
- 物理的/論理的役割: これは、空間の特定点における植生バイオマスの瞬間的な変化率を表します。
- 使用理由: 植生の時間的進化をモデル化するため。
-
$s$:
- 数学的定義: 正の定数パラメータ。
- 物理的/論理的役割: 降雨や降雨のような、システムへの水の一定の流入を表します。これは水のソース項です。
- 使用理由: 連続的な外部からの水の供給をモデル化するため。直接水のプールに寄与するため、加算項です。
-
$-w$:
- 数学的定義: 線形項、水の密度の負の値。
- 物理的/論理的役割: 蒸発による水の損失率を表します。元の次元モデルでは、これは $LW$ であり、$L$ は蒸発率でした。無次元化後、$L$ は暗黙的に吸収されるか1に設定されます。これは水のシンク項です。
- 使用理由: 水の存在に比例した水の損失をモデル化するため。水がシステムから除去されるため、減算項です。
-
$-\frac{pwn^2}{(w+k)(n+1)}$ (最初の式):
- 数学的定義: $p, w, n, k$ を含む複雑な非線形項。
- 物理的/論理的役割: これは、植生が水を吸収する速度を表す重要な「二重飽和変換項」です。$p$ は変換効率です。$n^2$ は、協同効果や表面積の増加により、水の吸収が植生密度とともに線形以上に増加することを示唆しています。分母の $(w+k)$ と $(n+1)$ は飽和を導入します。水 $w$ または植生 $n$ が非常に高くなると、水の吸収速度は無限に増加するのではなく、最大値に近づきます。この項は、水が植生によって消費されるため、水の式から差し引かれます。
- 使用理由: 水利用可能性と植生の吸収能力の両方の飽和効果を含む、植生による水の吸収という現実的な生物学的現象を捉えるため。乗算は $p, w, n^2$ の影響を組み合わせます。$(w+k)$ と $(n+1)$ による除算は飽和効果を生み出し、値が高くなると速度が平坦になり、これは生物学における制限されたリソースまたは容量をモデル化するためによく使用される方法です。
-
$+d_1\Delta w$:
- 数学的定義: 拡散項、$d_1$ は正の定数、$\Delta$ はラプラシアン演算子($\Delta w = \frac{\partial^2 w}{\partial x_1^2} + \frac{\partial^2 w}{\partial x_2^2} + \dots$)です。
- 物理的/論理的役割: 拡散(例:土壌への浸透、横方向の流れ)による水の空間的な広がりを表します。水は高濃度域から低濃度域へ移動する傾向があり、空間的な違いを平滑化します。正の符号はこの平滑化効果を示します。
- 使用理由: 水が空間に分散する自然な傾向をモデル化するため。局所的な水の密度を増加または減少させる可能性のある正味のフラックスを表すため、加算項です。
-
$\frac{pwn^2}{(w+k)(n+1)}$ (2番目の式):
- 数学的定義: 最初の式と同じ非線形項。
- 物理的/論理的役割: この項は、吸収された水をバイオマスに変換することによって植生が成長する速度を表します。これは植生のソース項です。
- 使用理由: 水の吸収と植生の成長を直接結びつけるため。このプロセスは植生バイオマスを増加させるため、加算項です。
-
$-bn$:
- 数学的定義: 線形項、$b$ と $n$ の積。$b$ は正の定数です。
- 物理的/論理的役割: 植生バイオマスの自然な死または損失率を表します。これは植生のシンク項です。
- 使用理由: 植生の自然な衰退または死亡率をモデル化するため。植生がシステムから除去されるため、減算項です。
-
$+d_2\Delta n$:
- 数学的定義: 拡散項、$d_2$ は正の定数、$\Delta$ は $n$ に適用されるラプラシアン演算子です。
- 物理的/論理的役割: 植生の空間的な広がり(例:無性生殖、種子散布)を表します。植生は高密度域から低密度域へ広がる傾向があります。
- 使用理由: 植生が空間に自然に分散するのをモデル化するため。$d_1\Delta w$ と同じ理由で加算項です。
-
$\frac{\partial w}{\partial \nu} = \frac{\partial n}{\partial \nu} = 0, x \in \partial\Omega, t > 0$:
- 数学的定義: ノイマン境界条件、ここで $\frac{\partial}{\partial \nu}$ は法線方向の微分(境界に垂直な変化率)を表します。
- 物理的/論理的役割: これらの条件は、空間領域の境界での「ゼロフラックス」または「流れなし」の制約を課します。これは、水や植生バイオマスがその端を通過してシステムに入ったり出たりしないことを意味します。
- 使用理由: すべての相互作用が指定された領域内で発生する閉鎖システムを定義し、外部からの影響が境界ダイナミクスに影響を与えるのを防ぐため。
-
$(w_0(x, 0), n_0(x, 0)) \geq, \neq (0,0), x \in \Omega$:
- 数学的定義: 水と植生の密度の初期条件。これらは非負であり、どこでも恒等的にゼロではありません。
- 物理的/論理的役割: 初期時刻 $t=0$ における水と植生バイオマスの開始空間分布を指定します。ゼロでない条件は、ダイナミクスが展開するための初期「種子」があることを保証します。
- 使用理由: システムの進化のシミュレーションまたは解析の開始点を提供するため。
ステップごとの流れ
私たちの生態系領域内の特定の場所で共存する、水のごく小さな抽象的な区画と植生のごく小さなパッチを想像してください。それらの旅を数学的エンジンを通して追ってみましょう。
- 水の流入: まず、水の区画は、降雨に似た一定の安定した水の供給、$s$ を受け取ります。これが主な入力です。
- 水の損失(蒸発): 同時に、区画の水の一部は蒸発によって環境に失われます。これは $-w$ 項で表されます。水が多いほど、蒸発も多くなります。
- 植生の渇き(水の観点): 次に、この場所の植生パッチが区画から水を吸収し始めます。この吸収は複雑なプロセスです。植生の密度($n^2$)と効率係数($p$)によって駆動されますが、制限も受けます。水の区画が非常に湿って($w$ が高い)または植生パッチが非常に密集して($n$ が高い)も、吸収速度は無限に増加し続けるのではなく、分母の $(w+k)$ と $(n+1)$ 項のおかげで「飽和」または平坦化します。この吸収された水は、水の区画から除去されます。
- 水の拡散: これらの局所的な相互作用の後、区画の水はそのまま留まりません。それは拡散し、隣接地域に広がり、同時に隣接地域からの水もこの区画に拡散します。このプロセスは、拡散係数 $d_1$ とラプラシアン演算子 $\Delta w$ によって支配され、空間的な濃度の違いを平滑化する平滑化メカニズムのように機能します。
- 植生の成長(植生の観点): 次に、植生パッチに焦点を移しましょう。吸収された水と同じ水($\frac{pwn^2}{(w+k)(n+1)}$ 項)が、新しい植生バイオマスに変換されます。これは植生の主な成長メカニズムであり、その拡大を水の利用可能性とそれ自体の密度に直接結び付けます。この新しいバイオマスは、植生パッチに追加されます。
- 植生の損失(死亡): しかし、植生も自然な損失に直面します。植生バイオマスの一定の割合が、その現在の密度($bn$)に比例して、死滅したり他の要因で失われたりします。これは植生パッチから差し引かれます。
- 植生の拡散: 水と同様に、植生バイオマスも空間的に広がります。拡散係数 $d_2$ とラプラシアン演算子 $\Delta n$ によって駆動され、植生は低密度地域に分散し、新しい植生がこのパッチに隣接地域から広がります。
- 境界の封じ込め: 全生態系領域の端では、「ゼロフラックス」条件が課せられます。これは、水や植生バイオマスが境界を通過してシステムに入ったり出たりしないことを意味し、システムを効果的に自己完結型ユニットにします。
- 連続的な進化: これらのステップは逐次的に発生するのではなく、領域内のすべての単一の点で連続的かつ同時に発生します。水と植生の初期分布から開始して、局所的な反応(流入、蒸発、吸収、成長、損失)と空間拡散のこの複雑な相互作用は、生態系全体を進化させ、安定した均一な状態または複雑で動的な空間パターンにつながる可能性があります。
最適化ダイナミクス
この反応拡散システムの文脈では、「最適化ダイナミクス」とは、モデルがデータから学習したり、機械学習の意味で損失関数を最小化したりすることではなく、システムが時間とともにどのように進化し、安定した状態またはパターンに収束するかを指します。ここでの「学習」は、システムが自然な構成を見つける固有の傾向です。
-
平衡への進化: システムの主なダイナミクスは、水と植生の両方の変化率がゼロになる平衡点($\frac{\partial w}{\partial t} = 0$ および $\frac{\partial n}{\partial t} = 0$)に向かって進化することです。これらの平衡は、入力と出力、および反応と拡散プロセスが完全にバランスしている定常状態を表します。本論文は、いくつかのそのような点を特定しています。「境界平衡」(例:植生がなく水のみの状態)と「正の平衡」(水と植生の両方がある状態)です。
-
安定性と「損失地形」: 平衡点の安定性の「損失地形」の概念は、直感的にマッピングできます。平衡点が安定している場合、システムはわずかに摂動されると、その状態に戻る傾向があります。不安定な場合は、わずかな摂動でもシステムが離れる原因となります。この地形の形状は、非線形相互作用と拡散項によって決定されます。本論文は、平衡点周りの線形化されたシステムのヤコビ行列の固有値を調べることによって線形安定性解析を使用します。
- 均一な平衡点の場合、すべての固有値の実部が負であれば、それは安定です。固有値の実部が正であれば、それは不安定です。
-
チューリング不安定性:パターン形成: 主要なダイナミクスはチューリング不安定性であり、これは空間パターンがどのように出現するかです。これは、空間的に均一な平衡点が均一な摂動に対して安定している(摂動が均一であれば均一性に戻る)が、空間的に変動する摂動に対して不安定になる(特定の空間波長の摂動が増加する)場合に発生します。
- 拡散係数($d_1$ および $d_2$)はここで重要です。それらが十分に異なれば、特定の空間周波数に対して均一な状態を不安定にし、斑点、ストライプ、またはギャップのようなパターンが自発的に形成される可能性があります。これは、システムがより複雑で空間的に不均一な「最適」をその状態空間で見つけるようなものです。本論文は、線形化されたシステムの固有値(これには拡散項が含まれるようになった)が異なる空間波数に対してどのように振る舞うかを調べることによってこれを分析します。
-
分岐ダイナミクス:質的変化: 主要なパラメータ(例:水の拡散係数 $d_1$)が変化すると、システムは分岐を起こす可能性があります。これは、解の質的な性質が劇的に変化することを意味します。例えば、安定な均一な状態が突然安定な空間パターンを生み出したり、その逆が起こったりする可能性があります。
- 本論文は、新しい平衡解(しばしば不均一なパターン)が既存のものから分岐する定常状態分岐を調査します。これらの分岐の方向(超臨界または亜臨界)は、新しいパターンが安定でスムーズに現れるか(超臨界)、それとも不安定で突然現れるか(亜臨界)を決定し、ヒステリシスにつながる可能性があります。これは、分岐関数の高次の導関数によって決定されます。
-
反復状態更新(数値シミュレーション): 数学的解析は安定性と分岐に関する理論的洞察を提供しますが、システムの実際の反復状態更新は数値シミュレーションを通じて視覚化されます。偏微分方程式は、空間と時間を離散化することによって計算的に解かれます。各短い時間ステップで、$w$ と $n$ の値は、局所的な反応項と隣接点からの拡散に基づいて、すべての空間グリッドポイントで更新されます。この反復プロセスにより、研究者は空間パターンの出現と進化を観察でき、理論的予測を確認し、システムがその動的構成にどのように「収束」するかを例示できます。本論文の図はこれを鮮やかに示しており、初期のランダムノイズが異なるパラメータ設定の下で組織化された斑点、ストライプ、またはギャップパターンに進化する様子を示しています。
結果、限界、結論
実験設計とベースライン
本研究の理論的発見の実験的検証は、提案された植生-水システム(1.5)の数値シミュレーションを通じて行われました。主な目的は、安定性、不安定性、およびパターン形成に関する数学的主張を徹底的に証明することでした。著者らは、特定の理論的予測に合致するように設計された2つの異なるパラメータ構成を選択することにより、実験を設計しました。
最初のパラメータセットでは、局所的漸近安定性を実証するために、値は $s = 4.69$, $p = 8.19$, $k = 1.3$, $b = 3.8$, $d_1 = 6$, $d_2 = 0.105$ に設定されました。これらのパラメータは、定理2.2によれば、平衡点 $(w_1, n_1)$ の局所的漸近安定性を予測する条件 $z < \dot{z} < z_2$ を満たすように特別に選択されました。
逆に、チューリング不安定性を示すことを意図した2番目のパラメータセットでは、値は $s = 4.69$, $p = 8.19$, $k = 1.3$, $b = 3.8$, $d_1 = 6$, $d_2 = 0.05$ でした。この構成は、定理2.5(2)によれば、平衡点 $(w_1, n_1)$ のチューリング不安定状態につながるはずの条件 $d_2\lambda_1 < \frac{b}{n_1+1}$ および $d_1 > \bar{d_1}$ を満たすように選択されました。
シミュレーションは $[0,50] \times [0,50]$ の2次元空間領域で実行され、$[0,600]$ 秒の時間間隔で進化しました。これらのシミュレーションの初期条件には、空間的に均一なシステムにおける定常解 $(w_1, n_1)$ への小さなランダム摂動の導入が含まれていました。この実験設計における「犠牲者」は外部のベースラインモデルではなく、理論的予測そのものでした。中核メカニズムの機能の決定的な証拠は、数値シミュレーションが予測された安定または不安定な挙動とそれに続くパターン形成を正確に再現したという直接的な視覚的および定量的確認でした。
証拠が証明すること
数値シミュレーションは、中核的な数学的メカニズム、特に二重飽和項と拡散係数の役割が、理論的解析によって予測された通りに機能したという否定できない証拠を提供しました。
-
安定性と不安定性の検証:
- 局所的漸近安定性: 最初のパラメータセットでは、図1はシステムの進化をグラフで示しています。「u1値」(水の密度)と「u2値」(植生バイオマス)のプロットは、初期摂動にもかかわらず、システムが時間とともに空間領域全体で均一で安定した状態に迅速に収束することを示しています。これは定理2.2を直接検証しており、特定の条件下では平衡点 $(w_1, n_1)$ が確かに局所的に漸近的に安定であることを確認しています。
-
チューリング不安定性: 2番目のパラメータセットでは、図2は安定した挙動とは対照的です。ここでは、「u1値」と「u2値」のプロットは、均一な状態ではなく、複雑な空間パターンの出現と持続を明確に示しています。この決定的なパターン形成はチューリング不安定性の特徴であり、定理2.5(2)の予測の確固たる証拠を提供します。初期に摂動されたシステムは、構造化された不均一な状態に進化したことを証明し、不安定性の理論的条件が正確に特定されたことを示しています。
-
動的な植生パターン形成:
- 時間経過に伴う植生分布を示す図3は、パターン進化の魅力的なシーケンスを明らかにしています。ランダムな摂動から開始して、システムは最初に無秩序で不均一な一時的なパターンを示します。時間が経過するにつれて(例えば、$t=2$秒から$t=50$秒)、ストライプ状のパターンが出現し始めます。最終的に、$t=600$秒までに、これらは主要なギャップパターンに進化し、その後変化しません。初期の無秩序から安定した複雑なパターンへのこの動的な進行は、モデルが現実的な生態学的現象を捉える能力があることの説得力のある証拠を提供します。青(低水分)と赤(高水分)の領域の視覚的な区別は、資源の空間分布をさらに明確にします。
-
主要な生態学的パラメータの影響:
- 水の拡散係数 ($d_1$): 図4の最初の重要な要因は水の拡散係数 $d_1$ です。$d_1 = 3$ の場合、図4(a)に示すように、規則的な高密度スポットが出現します。これらの条件下では水の拡散は遅く、植生は自然の草原パッチのように小さな集中したパッチで成長します。$d_1$ が増加し、図4(b)のシナリオに移行すると、パターンは複雑な迷路状のストライプに進化し、高水/低植生と低水/高植生領域が交互に現れます。$d_1$ が5になると、ストライプは細長くなり、バンドとスポットの複雑な混合物を形成します。水の拡散の強化は植生を伸ばし、より分散させます。$d_1$ が6.2に設定されると、図4(d)に示すように、大きくてまばらなパッチが現れます。高水領域は広く広がり、植生を孤立した小さな領域に押しやり、植生密度を低下させます。このシーケンスは、水の移動性の増加($d_1$ が高い)が植生の分散を促進し、バイオマス密度を低下させることを厳密に証明しています。
-
降雨強度 ($s$): 図5は降雨率の影響を示しています。最初に、$s$ が上昇するにつれて(例:$s=3.69$ から $s=4$)、ストライプパターンが出現します。$s$ のさらなる増加(例:$s=4.5$ から $s=4.69$)は、ギャップパターンの形成につながります。この証拠は、降雨と植生成長との間に非線形な関係があることを証明しており、特定の範囲内のより高い降雨率がより高い植生密度をもたらしますが、異なるパターンタイプへの遷移も引き起こします。
- 環境ストレス/資源競争 ($p$): 図6は、パラメータ $p$(環境ストレスまたは資源競争に関連)がゆっくり上昇すると、植生パターンはストライプ状の構造を維持しながらも、より狭く、より集中するようになることを示しています。これは、ストレスまたは競争の増加が、よりコンパクトで広がりの少ない植生分布につながることを示しています。
要するに、数値シミュレーションは、安定性と不安定性の理論的予測を検証しただけでなく、動的なパターン形成の豊かな視覚的タペストリーを提供しました。このハードな証拠は、主要なパラメータを体系的に変化させることから得られたものであり、モデルの中核メカニズム、二重飽和項の組み込みが、砂漠化しやすい環境で観察される複雑な生態水文学的フィードバックを正確に捉えていることを決定的に証明しました。
限界と将来の方向性
この研究は、新しい植生-水モデルの厳密で洞察に富んだ分析を提示していますが、その固有の限界を認識し、将来の開発への道筋を検討することが重要です。
即時の限界の1つは、二重飽和項によって導入された非線形性の増加に起因します。著者らが指摘するように、これは特に定常状態解の導出と分岐解析において、解析の複雑さを大幅に増大させます。本論文はこれらの課題をうまく乗り越えていますが、計算集約性と数学的な複雑さは、より広範な応用や、さらに複雑な生態学的相互作用の組み込みの障壁となる可能性があります。将来の研究では、このような高次の非線形性を効率的に処理するための高度な数値技術や機械学習アプローチを探求し、シミュレーションの高速化とより広範なパラメータ空間の探索を可能にする可能性があります。
数値シミュレーションの範囲も議論のポイントです。実験は、特定の2次元空間領域($[0,50] \times [0,50]$)と有限の時間間隔($[0,600]$)で実施されました。理論的主張を検証するには十分でしたが、これらの発見の、より大きな地理的スケール、より長い生態学的時間スケール、または異なる境界条件への一般化可能性は未解決の問題のままです。将来の研究では、これらのシミュレーションを地域または大陸レベルにスケールアップすることが含まれる可能性があり、地理情報システム(GIS)データと統合することで、現実世界の異質な景観におけるモデルの予測能力を評価できます。
さらに、モデルは、高度であるにもかかわらず、主に植生-水相互作用に焦点を当てています。砂漠化に影響を与える他の重要な生態学的要因、例えば土壌の種類、地形、家畜の放牧圧、火災レジーム、または異なる植物種(例:草対低木)の影響などは、モデルの中核方程式に明示的に詳細化されていません。これらの要素を組み込むことで、砂漠化ダイナミクスのさらに包括的で現実的な表現が可能になる可能性があります。例えば、土壌水保持が空間的に変動した場合、または放牧圧が動的変数として導入された場合、観察されたパターンはどう変化するでしょうか?これは、多種植生モデルや空間的に明示的な環境異質性を持つモデルにつながる可能性があります。
シミュレーションの初期条件には「小さなランダム摂動」が含まれていました。より大きな、またはより構造化された摂動下でのシステムの挙動を調査することは価値があるでしょう。これは、現実世界の擾乱(極端な気象イベントや人間の介入など)をよりよく模倣する可能性があります。これにより、小さな摂動では捉えられない異なる過渡ダイナミクスや代替安定状態が明らかになる可能性があります。
将来に向けて、これらの発見からいくつかの議論のトピックが浮上します。
- 予測モデリングと早期警報システム: モデルが観察された植生ダイナミクスを反映した多様な空間パターンを生成する能力を考えると、これを砂漠化のための予測ツールの開発にどのように活用できるでしょうか?特定のパターン遷移(例:斑点からストライプ、そしてギャップへ)は、半乾燥地域における生態系劣化の早期警報指標として機能するでしょうか?これには、長期的な生態学的モニタリングデータとの広範なキャリブレーションが必要になります。
- 資源管理の最適化: 水の拡散 ($d_1$) と降雨 ($s$) が植生パターンにどのように影響するかについての洞察は重要です。これらの発見は、水不足地域における持続可能な水配分戦略にどのように情報を提供できるでしょうか?例えば、まばらなパッチにつながる重要な $d_1$ 値を理解することは、土壌水保持を強化するか流出を減らすための措置を実施する land managers を導く可能性があり、それによってより回復力のある植生構造を促進します。
- 気候変動シナリオの影響: 単純な降雨強度の変化を超えて、このモデルは、干ばつの頻度と強度の増加、季節的な降雨パターンの変化、または蒸発率に影響を与える気温上昇のような、より複雑な気候変動予測にどのように応答するでしょうか?気候モデル出力をこのフレームワークに統合することで、将来の砂漠化リスクに関する貴重な洞察が得られる可能性があります。
- 学際的協力: これらの発見を真に進化させるためには、野外生態学者、水文学者、リモートセンシング専門家との緊密な協力が不可欠です。さまざまな砂漠化しやすい地域からの経験的データに対するモデルの予測を検証することは、その生態学的関連性と実用性を強化します。これには、リモートセンシングデータを使用して植生パターンと水利用可能性を推測することも含まれる可能性があり、その後、モデルパラメータのキャリブレーションと洗練に使用できます。
- 確率性と不確実性: 本論文は、確率性を組み込んだ以前の研究 [23] に言及しています。自然システムにおける固有の不確実性と変動性を反映した、確率的な環境変動下で二重飽和モデルはどのように振る舞うでしょうか?これは、予測不可能な条件下での植生の回復力に関するより堅牢な理解を提供する可能性があります。
- 高次元における分岐解析: 本論文は局所的および大域的な分岐解析を深く掘り下げていますが、モデルの複雑さにより、高次元空間設定におけるすべての分岐現象の完全な解析的治療は依然として大きな課題です。この分野におけるさらなる理論的進歩は、モデルの豊かなダイナミクスのより深い理解を解き放つ可能性があります。
これらの限界に対処し、これらの将来の方向性を探求することにより、本論文で提示された基礎的な作業は大幅に拡張され、砂漠化と戦い、生態系の回復力を促進するためのより効果的な戦略に貢献できます。
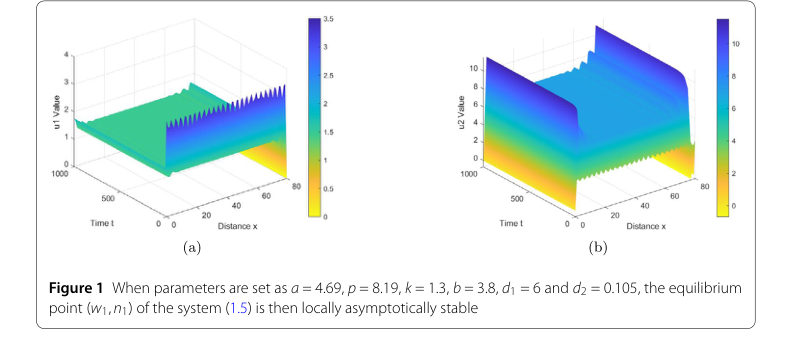 Figure 1. When parameters are set as a = 4.69, p = 8.19, k = 1.3, b = 3.8, d1 = 6 and d2 = 0.105, the equilibrium point (w1,n1) of the system (1.5) is then locally asymptotically stable
Figure 1. When parameters are set as a = 4.69, p = 8.19, k = 1.3, b = 3.8, d1 = 6 and d2 = 0.105, the equilibrium point (w1,n1) of the system (1.5) is then locally asymptotically stable
 Figure 2. When parameters are set as a = 4.69, p = 8.19, k = 1.3, b = 3.8, d1 = 6 and d2 = 0.05, the equilibrium point (w1,n1) of system (1.5) is Turing unstable
Figure 2. When parameters are set as a = 4.69, p = 8.19, k = 1.3, b = 3.8, d1 = 6 and d2 = 0.05, the equilibrium point (w1,n1) of system (1.5) is Turing unstable
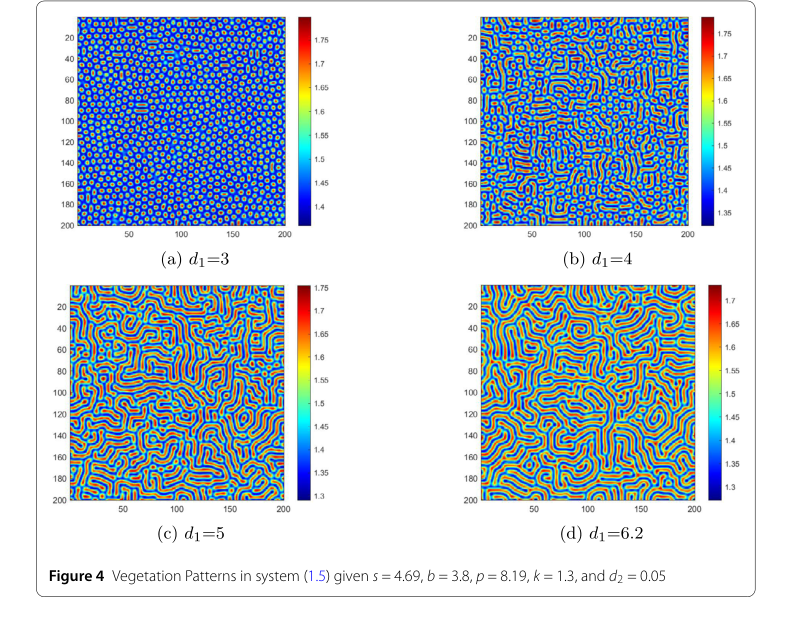 Figure 4. Vegetation Patterns in system (1.5) given s = 4.69, b = 3.8, p = 8.19, k = 1.3, and d2 = 0.05
Figure 4. Vegetation Patterns in system (1.5) given s = 4.69, b = 3.8, p = 8.19, k = 1.3, and d2 = 0.05