Динамическое поведение модели растительности и воды с членом двойного насыщения
Проблема опустынивания земель, являющаяся важнейшей экологической проблемой, широко распространенной в засушливых, полузасушливых и полувлажных регионах, составляет точное происхождение данного исследования.
Предпосылки и академическая родословная
Истоки и академическая родословная
Проблема опустынивания земель, являющаяся важнейшей экологической проблемой, широко распространенной в засушливых, полузасушливых и полувлажных регионах, составляет точное происхождение данного исследования. Она возникла в засушливых, полузасушливых и полувлажных регионах по всему миру из-за природных факторов и деятельности человека, ускоряющих опустынивание, что приводит к эрозии почв, сокращению растительности и нарушению стабильности экосистем. Академическая область математической экологии стремится моделировать эти сложные взаимодействия.
Исторически Клаусмайер [8] представил фундаментальную систему растительность-вода (модель 1.1), которая включала пространственную диффузию и перенос. Эта модель послужила основой для последующих исследований. Например, Шерратт и др. [9] интегрировали импульсную структуру для изучения спорадических осадков, а Консоло и др. [10] анализировали паттерны Тьюринга, используя обобщенную гиперболическую модель Клаусмайера. Кили и др. [11] упростили модель Клаусмайера (приведя к модели 1.2), удалив конвективные члены и используя линейную диффузию, что сделало ее более применимой к плоским ландшафтам. Эта упрощенная модель была далее исследована Сунь и др. [12], Чжан и др. [13] и Гоу и др. [14] для характеристики стабильных конфигураций, параметров бифуркации и влияния параметров на распределение растительности. Затем Ли и др. [29] ввели эффект насыщения водопоглощения в систему (модель 1.3), признавая, что растения физиологически регулируют водопоглощение, достигая максимальной поглощающей способности при определенном пороге биомассы.
Фундаментальным ограничением, или "болевой точкой", этих предыдущих подходов, включая модель (1.3) с ее единственным членом насыщения, была их неспособность полностью уловить двунаправленные взаимодействия насыщения между растительностью и водой. В то время как модель (1.3) учитывала насыщенное водопоглощение растительностью, она не моделировала явно характеристики насыщения самой доступности воды. Это упущение означало, что более ранние модели давали менее яркое и биологически точное изображение взаимного влияния и трансформационных отношений между растительностью и водой, особенно в регионах, подверженных опустыниванию, где и вода, и растительность могут одновременно становиться лимитирующими факторами. Этот пробел вынудил авторов разработать модель текущей статьи, которая включает "член двойного насыщения" для решения этой критической двунаправленной обратной связи, тем самым повышая биологическую достоверность модели, несмотря на увеличение аналитической сложности.
Интуитивные термины предметной области
- Опустынивание земель: Представьте себе некогда пышный сад, который медленно превращается в бесплодный, песчаный участок. Это происходит, когда почва теряет способность поддерживать растительную жизнь, часто из-за сочетания естественной засухи и деятельности человека, такой как чрезмерный выпас скота или вырубка лесов. Это похоже на то, как сад медленно "высыхает" и становится пустыней.
- Неустойчивость Тьюринга: Подумайте о ровном, однородно окрашенном тесте для торта. Если вы добавите определенные ингредиенты (например, определенные химикаты в реакции или скорости диффузии в биологической системе) и позволите им взаимодействовать, тесто может спонтанно развить четкие, повторяющиеся узоры — такие как полосы, пятна или лабиринты — по мере застывания. Неустойчивость Тьюринга описывает это явление, когда однородное состояние становится нестабильным и порождает сложные, организованные узоры.
- Член двойного насыщения: Рассмотрите голодное растение (растительность), пытающееся пить из источника воды. Во-первых, само растение может поглотить лишь определенное количество воды, независимо от того, сколько ее доступно (насыщение растительности). Во-вторых, если в почве изначально очень мало воды, растение не сможет поглотить много, даже если способно поглотить больше (насыщение воды). Этот член "двойного насыщения" в модели учитывает оба этих ограничения одновременно, делая взаимодействие более реалистичным.
- Бифуркационные явления: Представьте себе реку, текущую одним, устойчивым потоком. Если вы постепенно измените условие, такое как уклон местности или количество протекающей воды, река может внезапно разделиться на два отдельных русла, начать сильно извиваться или даже высохнуть. В математике бифуркация — это когда небольшое, непрерывное изменение параметра системы приводит к внезапному, качественному изменению ее поведения или количества и типа ее решений.
Таблица обозначений
| Обозначение | Описание | Тип |
|---|---|---|
| $w$ | Безразмерная плотность воды | Переменная |
| $n$ | Безразмерная биомасса растительности | Переменная |
| $x$ | Безразмерная пространственная координата | Переменная |
| $t$ | Безразмерное время | Переменная |
| $s$ | Безразмерный сток воды (интенсивность осадков) | Параметр |
| $p$ | Безразмерный параметр, связанный с ростом растительности/преобразованием воды | Параметр |
| $b$ | Безразмерный коэффициент убыли растительности | Параметр |
| $k$ | Безразмерный порог насыщения воды | Параметр |
| $d_1$ | Безразмерный коэффициент диффузии воды в почве | Параметр |
| $d_2$ | Безразмерный коэффициент диффузии растительности | Параметр |
Определение проблемы и ограничения
Основная постановка задачи и дилемма
Основная проблема, рассматриваемая в данной статье, заключается в разработке более биологически достоверной математической модели взаимодействия растительности и воды в регионах, подверженных опустыниванию. Предыдущие модели, такие как модель Клаусмайера (1.1) и ее расширения, такие как модель Кили и др. (1.2) и модель Ли и др. (1.3), учитывали эффект насыщения водопоглощающей способности растительности, представленный членом вида $F(N) = \frac{aN^2}{1+bN}$.
Входное/текущее состояние — это набор существующих моделей растительности и воды, которые, будучи фундаментальными, не полностью учитывают двунаправленные взаимодействия насыщения между растительностью и водой. В частности, им не хватает механизма для учета насыщения самой доступности воды, что приводит к менее реалистичному изображению экогидрологической обратной связи.
Выходное/целевое состояние — это новая система растительность-вода, в частности модель (1.4) и ее безразмерная форма (1.5), которая включает члены двойного насыщения. Эта новая модель призвана более точно и ярко описывать взаимное влияние и трансформационные отношения между растительностью и водой, тем самым повышая ее биологическую достоверность в представлении влажностно-растительных взаимодействий в районах, подверженных опустыниванию. Конечная цель — обеспечить более глубокий и реалистичный анализ динамического поведения, включая глобальную устойчивость, неустойчивость Тьюринга и различные бифуркационные явления, при этих уточненных условиях.
Точное недостающее звено или математический пробел, который данная статья пытается преодолеть, заключается в динамике насыщения воды. В то время как предыдущие модели учитывали насыщение водопоглощения растительностью ($F(N)$), они явно не моделировали насыщение самой воды. В данной статье вводится функция насыщения воды $G(W) = \frac{W}{W+K}$ (где $W$ — плотность воды, а $K$ — константа насыщения). Ключевым нововведением является комбинированный член двойного насыщения $RG(W) \cdot F(N) \cdot N$ (или его безразмерный эквивалент $\frac{pwn^2}{(w+k)(n+1)}$). Этот член точно характеризует синергетическое ограничение насыщенной водопоглощающей способности растительности и собственных характеристик насыщения воды, обеспечивая более точное объяснение нелинейной экогидрологической обратной связи.
Болезненный компромисс или дилемма, которая поймала предыдущих исследователей и на которую явно указывают авторы, — это баланс между биологической реалистичностью и аналитической разрешимостью. В статье говорится: "Включение функции насыщения воды G(W) существенно повышает биологическую достоверность модели, позволяя ей более реалистично описывать влажностно-растительные взаимодействия в районах, подверженных опустыниванию. Однако это также увеличивает аналитическую сложность, особенно при выводе стационарных решений и проведении анализа бифуркаций, из-за введения нелинейностей более высокого порядка." Это означает, что достижение более точного биологического представления достигается ценой значительного увеличения математической сложности решения и анализа системы.
Ограничения и режимы отказа
Проблема точного моделирования динамики растительности и воды в регионах, подверженных опустыниванию, чрезвычайно сложна из-за нескольких суровых, реалистичных стен, с которыми сталкиваются авторы:
-
Физические и экологические ограничения:
- Неотрицательные решения: Для любой биологической модели плотности воды ($w$) и растительности ($n$) должны оставаться неотрицательными. Система (1.5) подчиняется неотрицательным начальным данным $(w_0(x,0), n_0(x,0)) \ge (0,0)$, что является фундаментальным требованием для физической реалистичности. Неспособность поддерживать неотрицательность сделает модель биологически бессмысленной.
- Ограниченная область и граничные условия нулевого потока: Анализ проводится на ограниченной пространственной области $\Omega$ с гладкой границей, при условиях нулевого потока (Неймана) ($\frac{\partial w}{\partial \nu} = \frac{\partial n}{\partial \nu} = 0$ на $\partial \Omega$). Эти условия представляют собой замкнутую систему, где вода или растительность не пересекают границы, что является реалистичным предположением для многих экологических участков, но добавляет сложности к дифференциальным уравнениям в частных производных (ДУЧП).
- Явления насыщения: Суть проблемы заключается в точном улавливании насыщения как водопоглощения растительностью, так и самой доступности воды. Это сложная физиологическая и экологическая реальность, которую необходимо встроить в модель.
- Факторы окружающей среды: Модель должна неявно или явно учитывать различные природные факторы (осадки, скорость ветра, свет) и деятельность человека (животноводство), которые влияют на распределение растительности и динамику воды.
-
Вычислительные и математические ограничения:
- Нелинейности высокого порядка: Введение члена двойного насыщения $G(W) \cdot F(N)$ значительно увеличивает нелинейность системы (1.4) и ее безразмерной формы (1.5). Это делает вывод стационарных решений, анализ устойчивости и анализ бифуркаций существенно более сложными по сравнению с линейными или нелинейными системами низкого порядка. Стандартные аналитические методы часто испытывают трудности с такой высокой степенью нелинейности.
- Аналитическая сложность для стационарных решений: Нахождение явных стационарных решений для сильно нелинейных систем реакция-диффузия, как правило, очень затруднительно. Статья опирается на строгие математические выводы, априорные оценки и качественные анализы, которые являются вычислительно интенсивными и требуют продвинутых математических инструментов.
- Проблемы анализа бифуркаций: Анализ бифуркационных явлений, особенно связанных с простыми и двойными собственными значениями, требует сложных методов, таких как метод редукции Ляпунова-Шмидта и теорема Крэндалла-Рабиновица. Эти методы сложны, и их применение становится более трудоемким при увеличении нелинейности. В статье отмечается, что при определенных условиях (например, когда $j \neq 2i$ и $i \neq 2j$) теорема о неявной функции не может быть применена для установления существования решений, что указывает на режим отказа для стандартных аналитических подходов.
- Условия неустойчивости Тьюринга: Определение условий, при которых возникает неустойчивость Тьюринга (ведущая к образованию паттернов), включает сложный анализ собственных значений линеаризованной системы, который сильно зависит от коэффициентов диффузии и других параметров.
- Исследование пространства параметров: Модель включает в себя многочисленные параметры (например, $s, p, b, k, d_1, d_2$). Тщательное исследование влияния каждого параметра на динамическое поведение системы и образование паттернов требует обширного теоретического анализа и численных симуляций, что делает пространство проблемы обширным и трудным для полного исследования. Численные симуляции в статье необходимы для проверки теоретических предсказаний в этом сложном ландшафте параметров.
Почему такой подход
Неизбежность выбора
Авторы данной статьи столкнулись с фундаментальным ограничением существующих моделей растительности и воды: они не смогли адекватно уловить сложные, двунаправленные эффекты насыщения между растительностью и водой в регионах, подверженных опустыниванию. Традиционные модели, такие как модель Клаусмайера (система 1.1) и ее расширения (например, система 1.3 Ли и др.), в основном включали единственный член насыщения, обычно описывающий, как водопоглощающая способность растительности насыщается по мере увеличения ее биомассы, представленный функцией вида $f(N) = \frac{aN^2}{1+bN}$.
Однако авторы осознали, что этого недостаточно. Точный момент этого осознания сформулирован на странице 3, где они заявляют: "Чтобы лучше уловить двунаправленные взаимодействия насыщения между растительностью и водой в регионах, подверженных опустыниванию, мы далее уточняем систему (1.3), включая механизм двойного насыщения." Это подчеркивает, что проблема заключается не только в поглощении воды растительностью, но и в собственных характеристиках насыщения воды и том, как это одновременно ограничивает рост растительности. Без учета обоих факторов модели упускали критически важную часть экологической головоломки. Следовательно, модель, включающая члены двойного насыщения, была не просто улучшением, а единственным жизнеспособным решением для точного представления этих сложных, взаимных ограничений. Это был явный недостаток.
Сравнительное превосходство
Качественное превосходство данного подхода проистекает непосредственно из его повышенной биологической достоверности, а не просто из улучшенных метрик производительности в вычислительном смысле. Введение "членов двойного насыщения" посредством нового нелинейного связующего члена $RG(W) \cdot F(N) \cdot N$ является ключевым структурным преимуществом. Этот член, как объясняется на странице 3, "характеризует одновременное ограничение воды и растительности в динамике поглощения".
Предыдущие модели, учитывая только одинарное насыщение, не могли полностью представить это синергетическое ограничение. Включение функции насыщения воды $G(W)$ наряду со скоростью поглощения растительностью $F(N)$ позволяет модели "более реалистично описывать влажностно-растительные взаимодействия в районах, подверженных опустыниванию" и "более точно объяснять нелинейный механизм экогидрологической обратной связи". Это означает, что модель не просто лучше подгоняет данные; она структурно разработана для более точного отражения реальных экологических процессов. Она обеспечивает более глубокое, более тонкое понимание того, как эти два критических компонента взаимодействуют в условиях стресса, что является значительным качественным скачком по сравнению с предыдущими золотыми стандартами. В статье не обсуждаются такие аспекты, как обработка высокоразмерного шума или снижение сложности памяти, поскольку основное внимание уделяется фундаментальному математическому моделированию экологических явлений.
Соответствие ограничениям
Честно говоря, я не совсем уверен в конкретных ограничениях, определенных в Шаге 2, поскольку этот раздел не был предоставлен. Однако, основываясь на определении проблемы и мотивации, представленных в статье, мы можем вывести несколько суровых требований, которым выбранный метод идеально соответствует. Основная проблема заключается в точном моделировании взаимодействий растительности и воды в "регионах, подверженных опустыниванию" (Аннотация, стр. 3), что подразумевает необходимость высокой достоверности в улавливании критических механизмов обратной связи в условиях экологического стресса.
"Связь" между требованиями проблемы и уникальными свойствами решения очевидна в нескольких аспектах:
1. Реалистичное изображение взаимного влияния: Проблема требует модели, которая "более ярко изображает взаимное влияние и трансформационные отношения между растительностью и водой" (Аннотация). Члены двойного насыщения, $G(W)$ и $F(N)$, напрямую удовлетворяют этому, явно моделируя, как и доступность воды, и поглощающая способность растительности одновременно ограничивают друг друга.
2. Двунаправленные взаимодействия насыщения: Авторы явно стремились "лучше уловить двунаправленные взаимодействия насыщения между растительностью и водой" (стр. 3). Новый член двойного насыщения $RG(W) \cdot F(N) \cdot N$ разработан именно для этого, гарантируя, что модель учитывает насыщение с обеих сторон взаимодействия, что было критическим упущением в предыдущих моделях.
3. Объяснение нелинейных механизмов: Опустынивание включает сложные, нелинейные экогидрологические обратные связи. Выбранный метод, с его нелинейностями более высокого порядка, введенными членами двойного насыщения, позволяет "более точно объяснять нелинейный механизм экогидрологической обратной связи в районах, подверженных опустыниванию" (стр. 3). Это идеально соответствует необходимости понимания сложных динамик, которые приводят к образованию паттернов и стабильности экосистем в этих уязвимых средах.
Отклонение альтернатив
Причина отклонения альтернативных подходов авторами в основном неявная и сосредоточена на ограничениях предыдущих математических моделей, а не на совершенно иных методологических парадигмах, таких как GANs или Трансформеры. Авторы опираются на ряд моделей реакция-диффузия, начиная с системы Клаусмайера (1.1) и ее последующих расширений (например, системы Кили и др. (1.2), системы Ли и др. (1.3)).
Основная причина отклонения этих более ранних моделей как достаточных заключается в их неспособности уловить "двунаправленные взаимодействия насыщения между растительностью и водой" (стр. 3). В то время как эти модели включали единственный член насыщения (например, $f(N) = \frac{aN^2}{1+bN}$ для водопоглощения растительностью), им не хватало соответствующего члена для насыщения воды, $G(W)$. Авторы обнаружили, что "традиционные" методы, в данном контексте означающие модели растительность-вода с одинарным насыщением, были "недостаточны для данной конкретной проблемы", поскольку они не могли "одновременно уловить эффекты насыщения как водопоглощающей способности растительности, так и доступности воды" (стр. 4). Это упущение привело к менее реалистичному изображению экогидрологической обратной связи. Следовательно, новая модель с ее членами двойного насыщения была представлена как необходимое уточнение для преодоления этих конкретных недостатков ее предшественников в рамках той же системы моделирования. В статье не рассматривается, почему модели машинного обучения, такие как GANs или модели диффузии, потерпели бы неудачу, поскольку они работают на другом уровне абстракции и не могут быть напрямую сопоставлены с подходом механистического моделирования дифференциальных уравнений, принятым здесь.
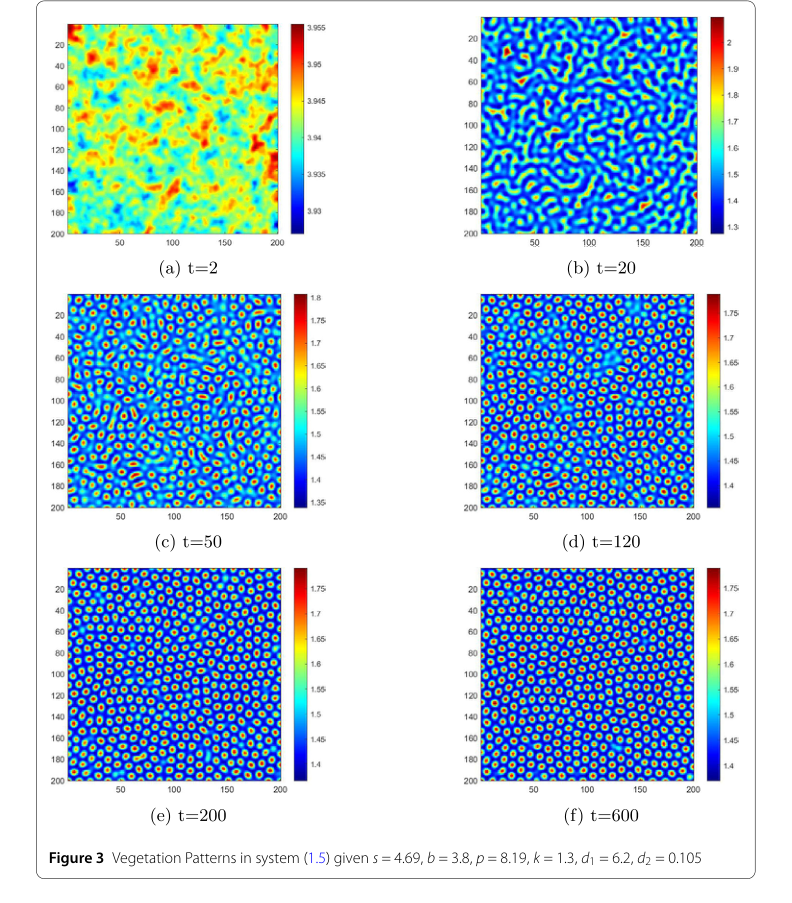 Figure 3. Vegetation Patterns in system (1.5) given s = 4.69, b = 3.8, p = 8.19, k = 1.3, d1 = 6.2, d2 = 0.105
Figure 3. Vegetation Patterns in system (1.5) given s = 4.69, b = 3.8, p = 8.19, k = 1.3, d1 = 6.2, d2 = 0.105
Математический и логический механизм
Мастер-уравнение
Абсолютным ядром математического механизма данной статьи является безразмерная система реакция-диффузия, которая описывает пространственно-временную эволюцию и формирование паттернов взаимодействия между биомассой растительности и плотностью воды. Эта система, выведенная из исходной модели путем включения членов двойного насыщения и последующего масштабирования, представлена следующим образом:
$$ \begin{cases} \frac{\partial w}{\partial t} = s - w - \frac{pwn^2}{(w+k)(n+1)} + d_1\Delta w, & x \in \Omega, t > 0, \\ \frac{\partial n}{\partial t} = \frac{pwn^2}{(w+k)(n+1)} - bn + d_2\Delta n, & x \in \Omega, t > 0, \\ \frac{\partial w}{\partial \nu} = \frac{\partial n}{\partial \nu} = 0, & x \in \partial\Omega, t > 0, \\ (w_0(x, 0), n_0(x, 0)) \geq, \neq (0,0), & x \in \Omega. \end{cases} $$
Поэлементный разбор
Давайте разберем это мастер-уравнение по частям, чтобы понять его компоненты и их роли:
-
$w$:
- Математическое определение: Зависимая переменная, представляющая безразмерную плотность воды. Это скалярное поле, что означает, что его значение может варьироваться в пространстве и времени.
- Физическая/логическая роль: Этот член количественно определяет количество воды, доступной в экосистеме в любой момент времени $t$ и в любой точке $x$.
- Почему используется: Это основная переменная состояния для воды.
-
$n$:
- Математическое определение: Зависимая переменная, представляющая безразмерную плотность биомассы растительности. Как и $w$, это скалярное поле.
- Физическая/логическая роль: Этот член количественно определяет количество биомассы растительности, присутствующей в любой момент времени $t$ и в любой точке $x$.
- Почему используется: Это основная переменная состояния для растительности.
-
$t$:
- Математическое определение: Независимая переменная, представляющая время.
- Физическая/логическая роль: Это временное измерение, по которому эволюционирует система.
- Почему используется: Для описания того, как $w$ и $n$ изменяются со временем.
-
$x \in \Omega, t > 0$:
- Математическое определение: Определяет область для дифференциальных уравнений в частных производных (ДУЧП). $\Omega$ — ограниченная пространственная область, а $t > 0$ означает, что мы рассматриваем эволюцию системы после начального момента.
- Физическая/логическая роль: Определяет физическое пространство и временной интервал, к которым применима модель.
- Почему используется: Для установки контекста пространственной и временной динамики.
-
$\frac{\partial w}{\partial t}$:
- Математическое определение: Частная производная плотности воды $w$ по времени $t$.
- Физическая/логическая роль: Это представляет собой мгновенную скорость изменения плотности воды в конкретной точке пространства. Положительное значение означает увеличение количества воды, отрицательное — уменьшение.
- Почему используется: Для моделирования временной эволюции воды.
-
$\frac{\partial n}{\partial t}$:
- Математическое определение: Частная производная плотности биомассы растительности $n$ по времени $t$.
- Физическая/логическая роль: Это представляет собой мгновенную скорость изменения биомассы растительности в конкретной точке пространства.
- Почему используется: Для моделирования временной эволюции растительности.
-
$s$:
- Математическое определение: Положительный постоянный параметр.
- Физическая/логическая роль: Представляет собой постоянный приток воды в систему, аналогичный осадкам или дождю. Это источник для воды.
- Почему используется: Для моделирования непрерывного внешнего снабжения водой. Это аддитивный член, поскольку он напрямую вносит вклад в водный пул.
-
$-w$:
- Математическое определение: Линейный член, отрицательное значение плотности воды.
- Физическая/логическая роль: Представляет собой скорость потери воды из-за испарения. В исходной размерной модели это было $LW$, где $L$ — скорость испарения. После безразмеривания $L$ неявно поглощается или устанавливается равной 1. Это стоковый член для воды.
- Почему используется: Для моделирования потери воды, пропорциональной ее наличию. Это вычитаемый член, поскольку вода удаляется из системы.
-
$-\frac{pwn^2}{(w+k)(n+1)}$ (в первом уравнении):
- Математическое определение: Сложный нелинейный член, включающий $p, w, n, k$.
- Физическая/логическая роль: Это критический "член двойного насыщения", который представляет собой скорость, с которой растительность поглощает воду. $p$ — это эффективность преобразования. $n^2$ предполагает, что водопоглощение увеличивается более чем линейно с плотностью растительности, возможно, из-за кооперативных эффектов или увеличения площади поверхности. Знаменатели $(w+k)$ и $(n+1)$ вводят насыщение: когда вода $w$ или растительность $n$ становятся очень высокими, скорость водопоглощения не увеличивается бесконечно, а приближается к максимуму. Этот член вычитается из уравнения воды, поскольку вода потребляется растительностью.
- Почему используется: Для улавливания реалистичного биологического явления водопоглощения растительностью, включая эффекты насыщения как доступности воды, так и поглощающей способности растительности. Умножение объединяет влияния $p, w, n^2$. Деление на $(w+k)$ и $(n+1)$ создает эффект насыщения, где скорость выравнивается при высоких значениях, что является распространенным способом моделирования ограниченных ресурсов или мощностей в биологии.
-
$+d_1\Delta w$:
- Математическое определение: Член диффузии, где $d_1$ — положительная константа, а $\Delta$ — оператор Лапласа ($\Delta w = \frac{\partial^2 w}{\partial x_1^2} + \frac{\partial^2 w}{\partial x_2^2} + \dots$).
- Физическая/логическая роль: Представляет собой пространственное распространение воды за счет диффузии (например, инфильтрация через почву, боковой поток). Вода имеет тенденцию перемещаться из областей с более высокой концентрацией в области с более низкой концентрацией, сглаживая пространственные различия. Положительный знак указывает на этот сглаживающий эффект.
- Почему используется: Для моделирования естественной тенденции воды к рассеиванию в пространстве. Это аддитивный член, поскольку он представляет собой чистый поток, который может увеличивать или уменьшать локальную плотность воды в зависимости от кривизны профиля воды.
-
$\frac{pwn^2}{(w+k)(n+1)}$ (во втором уравнении):
- Математическое определение: Идентичный нелинейный член, как и в первом уравнении.
- Физическая/логическая роль: Этот член представляет собой скорость, с которой растительность растет, преобразуя поглощенную воду в биомассу. Это источник для растительности.
- Почему используется: Для прямой связи водопоглощения с ростом растительности. Это аддитивный член, поскольку этот процесс увеличивает биомассу растительности.
-
$-bn$:
- Математическое определение: Линейный член, произведение $b$ и $n$. $b$ — положительная константа.
- Физическая/логическая роль: Представляет собой естественную скорость гибели или потери биомассы растительности. Это стоковый член для растительности.
- Почему используется: Для моделирования естественного увядания или смертности растительности. Это вычитаемый член, поскольку растительность удаляется из системы.
-
$+d_2\Delta n$:
- Математическое определение: Член диффузии, где $d_2$ — положительная константа, а $\Delta$ — оператор Лапласа, примененный к $n$.
- Физическая/логическая роль: Представляет собой пространственное распространение растительности (например, через бесполое размножение, распространение семян). Растительность имеет тенденцию распространяться из областей с более высокой плотностью в области с более низкой плотностью.
- Почему используется: Для моделирования естественного рассеивания растительности в пространстве. Это аддитивный член по тем же причинам, что и $d_1\Delta w$.
-
$\frac{\partial w}{\partial \nu} = \frac{\partial n}{\partial \nu} = 0, x \in \partial\Omega, t > 0$:
- Математическое определение: Граничные условия Неймана, где $\frac{\partial}{\partial \nu}$ обозначает нормальную производную (скорость изменения перпендикулярно границе).
- Физическая/логическая роль: Эти условия налагают ограничение "нулевого потока" или "отсутствия потока" на границах пространственной области. Это означает, что вода или биомасса растительности не входят и не покидают систему через ее края.
- Почему используется: Для определения замкнутой системы, где все взаимодействия происходят в пределах указанной области, предотвращая влияние внешних факторов на динамику границы.
-
$(w_0(x, 0), n_0(x, 0)) \geq, \neq (0,0), x \in \Omega$:
- Математическое определение: Начальные условия для плотности воды и растительности. Они неотрицательны и не равны нулю тождественно везде.
- Физическая/логическая роль: Задает начальное пространственное распределение воды и биомассы растительности в начальный момент времени $t=0$. Условие ненулевого значения гарантирует наличие начального "затравки" для развертывания динамики.
- Почему используется: Для предоставления отправной точки для симуляции или анализа эволюции системы.
Пошаговый поток
Представьте себе крошечный, абстрактный участок воды и небольшой участок растительности, сосуществующий в определенном месте в пределах нашей экологической области. Давайте проследим их путь через математический механизм:
- Приток воды: Во-первых, участок воды получает постоянный, стабильный приток воды, $s$, подобно осадкам. Это его основной вход.
- Потеря воды (испарение): Одновременно часть воды в участке теряется в окружающую среду из-за испарения, представленного членом $-w$. Чем больше воды, тем больше испаряется.
- Жажда растительности (с точки зрения воды): Далее, растительность в этом месте начинает поглощать воду из участка. Это поглощение — сложный процесс: он обусловлен плотностью растительности ($n^2$) и эффективностью ($p$), но также ограничен. Когда водный участок становится очень влажным (высокий $w$) или растительность становится очень плотной (высокий $n$), скорость поглощения не продолжает бесконечно увеличиваться; она "насыщается" или выравнивается благодаря членам $(w+k)$ и $(n+1)$ в знаменателе. Эта поглощенная вода удаляется из водного участка.
- Распространение воды: После этих локальных взаимодействий вода в участке не остается на месте. Она диффундирует, распространяясь в соседние области, в то время как вода из соседних областей также диффундирует в этот участок. Этот процесс, управляемый коэффициентом диффузии $d_1$ и оператором Лапласа $\Delta w$, действует как механизм сглаживания, стремясь выровнять концентрации воды в пространстве.
- Рост растительности (с точки зрения растительности): Теперь переключимся на участок растительности. Та же самая вода, которая была поглощена из участка (член $\frac{pwn^2}{(w+k)(n+1)}$), преобразуется в новую биомассу растительности. Это основной механизм роста растительности, напрямую связывающий ее расширение с доступностью воды и ее собственной плотностью. Эта новая биомасса добавляется к участку растительности.
- Потеря растительности (смертность): Однако растительность также подвержена естественным потерям. Часть биомассы растительности, пропорциональная ее текущей плотности ($bn$), увядает или теряется из-за других факторов. Это вычитается из участка растительности.
- Распространение растительности: Подобно воде, биомасса растительности также пространственно распространяется. Управляемая коэффициентом диффузии $d_2$ и оператором Лапласа $\Delta n$, растительность рассеивается в менее плотные области, и новая растительность распространяется в этот участок из соседних.
- Ограничение границ: На самых краях всей экологической области накладывается условие "нулевого потока". Это означает, что вода или биомасса растительности не могут входить или выходить из системы через ее границы, фактически делая систему самодостаточной единицей.
- Непрерывная эволюция: Эти шаги происходят не последовательно, а непрерывно и одновременно в каждой точке области. Со временем, начиная с начального распределения воды и растительности, это сложное взаимодействие локальных реакций (приток, испарение, поглощение, рост, потери) и пространственной диффузии вызывает эволюцию всей экосистемы, потенциально приводя к стабильным однородным состояниям или сложным, динамическим пространственным паттернам.
Динамика оптимизации
В контексте данной системы реакция-диффузия "динамика оптимизации" относится не к обучению модели на данных или минимизации функции потерь в смысле машинного обучения, а скорее к тому, как система эволюционирует и сходится к стабильным состояниям или паттернам со временем. "Обучение" здесь — это внутреннее стремление системы найти свои естественные конфигурации.
-
Эволюция к равновесиям: Основная динамика системы заключается в эволюции к точкам равновесия, где скорости изменения как воды, так и растительности становятся равными нулю ($\frac{\partial w}{\partial t} = 0$ и $\frac{\partial n}{\partial t} = 0$). Эти равновесия представляют собой стационарные состояния, где входы и выходы, а также процессы реакции и диффузии идеально сбалансированы. В статье выделено несколько таких точек: "граничное равновесие" (например, состояние только с водой и без растительности) и "положительные равновесия" (состояния с водой и растительностью).
-
Устойчивость и "ландшафт потерь": Концепция "ландшафта потерь" может быть интуитивно сопоставлена с ландшафтом устойчивости этих равновесий. Если равновесие устойчиво, система, будучи слегка возмущенной, будет стремиться вернуться в это состояние. Если оно неустойчиво, даже малейшее возмущение заставит систему отойти. Форма этого ландшафта определяется нелинейными взаимодействиями и членами диффузии. В статье используется линейный анализ устойчивости путем рассмотрения собственных значений якобиана линеаризованной системы вокруг равновесия.
- Для однородного равновесия, если все собственные значения имеют отрицательные действительные части, оно устойчиво. Если любое собственное значение имеет положительную действительную часть, оно неустойчиво.
-
Неустойчивость Тьюринга: Формирование паттернов: Ключевой динамикой является неустойчивость Тьюринга, которая определяет, как возникают пространственные паттерны. Это происходит, когда пространственно однородное равновесие устойчиво к однородным возмущениям (означает, что оно вернется к однородности при равномерном возмущении), но становится неустойчивым к пространственно изменяющимся возмущениям (означает, что возмущения определенной пространственной длины волны растут).
- Коэффициенты диффузии ($d_1$ и $d_2$) здесь критичны. Если они достаточно различны, они могут дестабилизировать однородное состояние для определенных пространственных частот, что приведет к спонтанному формированию паттернов, таких как пятна, полосы или пробелы. Это похоже на то, как система "находит" более сложный, пространственно неоднородный "оптимум" в своем пространстве состояний. В статье это анализируется путем рассмотрения того, как собственные значения линеаризованной системы (которые теперь включают члены диффузии) ведут себя для различных пространственных волновых чисел.
-
Динамика бифуркаций: Качественные сдвиги: По мере изменения ключевых параметров (например, коэффициента диффузии воды $d_1$) система может претерпевать бифуркации. Это означает, что качественная природа решений резко меняется. Например, стабильное однородное состояние может внезапно породить стабильные пространственные паттерны, или наоборот.
- В статье исследуются бифуркации стационарных состояний, где новые равновесные решения (часто неоднородные паттерны) ответвляются от существующих. Направление этих бифуркаций (сверхкритические или субкритические) определяет, являются ли новые паттерны стабильными и появляются ли они плавно (сверхкритические) или являются нестабильными и появляются внезапно (субкритические), что может привести к гистерезису. Это определяется производными более высокого порядка бифуркационной функции.
-
Итеративные обновления состояний (численное моделирование): В то время как математический анализ дает теоретическое понимание устойчивости и бифуркаций, фактические "итеративные обновления" состояния системы со временем визуализируются через численные симуляции. Дифференциальные уравнения в частных производных решаются вычислительно путем дискретизации пространства и времени. На каждом малом временном шаге значения $w$ и $n$ в каждой пространственной точке сетки обновляются на основе локальных членов реакции и диффузии из соседних точек. Этот итеративный процесс позволяет исследователям наблюдать возникновение и эволюцию пространственных паттернов, подтверждая теоретические предсказания и иллюстрируя, как система "сходится" к своим динамическим конфигурациям. Рисунки в статье наглядно демонстрируют это, показывая, как начальный случайный шум эволюционирует в организованные пятна, полосы или пробельные паттерны при различных параметрах.
Результаты, ограничения и заключение
Дизайн эксперимента и базовые уровни
Экспериментальная проверка теоретических выводов данного исследования проводилась посредством численных симуляций предложенной системы растительность-вода (1.5). Основная цель заключалась в безжалостной проверке математических утверждений относительно устойчивости, неустойчивости и формирования паттернов. Авторы спроектировали свои эксперименты, выбрав две различные конфигурации параметров, каждая из которых была предназначена для соответствия конкретным теоретическим предсказаниям.
Для первого набора параметров, выбранного для демонстрации локальной асимптотической устойчивости, значения были установлены как $s = 4.69$, $p = 8.19$, $k = 1.3$, $b = 3.8$, $d_1 = 6$ и $d_2 = 0.105$. Эти параметры были специально выбраны для удовлетворения условия $z < \dot{z} < z_2$, которое, согласно Теореме 2.2, предсказывает локальную асимптотическую устойчивость точки равновесия $(w_1, n_1)$.
Напротив, для второго набора параметров, предназначенного для иллюстрации неустойчивости Тьюринга, значения были $s = 4.69$, $p = 8.19$, $k = 1.3$, $b = 3.8$, $d_1 = 6$ и $d_2 = 0.05$. Эта конфигурация была выбрана для удовлетворения условий $d_2\lambda_1 < \frac{b}{n_1+1}$ и $d_1 > \bar{d_1}$, которые, согласно Теореме 2.5 (2), должны привести к неустойчивому состоянию Тьюринга точки равновесия $(w_1, n_1)$.
Симуляции проводились на двумерной пространственной области, определенной как $[0,50] \times [0,50]$, и эволюционировали в течение временного интервала $[0,600]$ секунд. Начальные условия для этих симуляций включали введение небольших случайных возмущений к стационарным решениям $(w_1, n_1)$ в пространственно однородной системе. "Жертвами" в этом экспериментальном дизайне были не внешние базовые модели, а сами теоретические предсказания. Окончательным доказательством работы основного механизма было прямое визуальное и количественное подтверждение того, что численные симуляции точно воспроизвели предсказанное стабильное или нестабильное поведение и последующее формирование паттернов.
Что доказывают свидетельства
Численные симуляции предоставили неоспоримые доказательства того, что основной математический механизм, в частности роль членов двойного насыщения и коэффициентов диффузии, работал так, как предсказывал теоретический анализ.
-
Проверка устойчивости и неустойчивости:
- Локальная асимптотическая устойчивость: Для первого набора параметров Рисунок 1 графически иллюстрирует эволюцию системы. Графики "Значение u1" (плотность воды) и "Значение u2" (биомасса растительности) показывают, что, несмотря на начальные возмущения, система быстро сходится к однородному, стабильному состоянию по всей пространственной области с течением времени. Это напрямую подтверждает Теорему 2.2, подтверждая, что при определенных условиях точка равновесия $(w_1, n_1)$ действительно локально асимптотически устойчива.
-
Неустойчивость Тьюринга: Со вторым набором параметров Рисунок 2 резко контрастирует со стабильным поведением. Здесь графики "Значение u1" и "Значение u2" явно показывают возникновение и сохранение сложных пространственных паттернов вместо однородного состояния. Это окончательное формирование паттернов является отличительной чертой неустойчивости Тьюринга, предоставляя убедительные доказательства теоретических условий неустойчивости, которые были точно определены. Система, изначально возмущенная, эволюционирует в структурированное, неоднородное состояние, доказывая, что теоретические условия неустойчивости были точно определены.
-
Динамическое формирование паттернов растительности:
- Рисунок 3, иллюстрирующий распределение растительности во времени, раскрывает захватывающую последовательность эволюции паттернов. Начиная со случайных возмущений, система изначально демонстрирует нерегулярные, неоднородные временные паттерны. По мере прогрессирования времени (например, от $t=2$ с до $t=50$ с) начинают появляться полосовидные паттерны. В конечном итоге, к $t=600$ с, они эволюционируют в доминирующие пробельные паттерны, которые затем остаются неизменными. Эта динамическая прогрессия от начального беспорядка к стабильным, сложным паттернам предоставляет убедительные доказательства способности модели улавливать реалистичные экологические явления. Визуальное различие между областями с низким содержанием влаги (синий) и высоким содержанием влаги (красный) далее поясняет пространственное распределение ресурсов.
-
Влияние ключевых экологических параметров:
- Коэффициент диффузии воды ($d_1$): На рис. 4 первый ключевой фактор — коэффициент диффузии воды $d_1$. При $d_1 = 3$, как показано на рис. 4 (а), возникают регулярные пятна с высокой плотностью. В этих условиях диффузия воды медленная, и растительность растет небольшими концентрированными участками, подобно травяным участкам в природе. По мере увеличения $d_1$, переходя к сценарию на рис. 4 (б), паттерн эволюционирует в сложные лабиринтоподобные полосы, чередующиеся области с высокой водой/низкой растительностью и низкой водой/высокой растительностью. Когда $d_1$ достигает 5, полосы удлиняются, образуя сложную смесь полос и пятен; усиленная диффузия воды растягивает растительность, делая ее более рассеянной. Когда $d_1$ устанавливается на уровне 6.2, как показано на рис. 4 (d), появляются большие, разреженные пятна. Области с высокой водой широко расширяются, вытесняя растительность в изолированные небольшие участки, вызывая снижение плотности растительности. Эта последовательность строго доказывает, что увеличение подвижности влаги (более высокое $d_1$) способствует рассеиванию растительности и снижает плотность биомассы.
-
Интенсивность осадков ($s$): Рисунок 5 иллюстрирует влияние интенсивности дождя. Изначально, при увеличении $s$ (например, от $s=3.69$ до $s=4$), появляются полосовидные паттерны. Дальнейшее увеличение $s$ (например, от $s=4.5$ до $s=4.69$) приводит к формированию пробельных паттернов. Эти доказательства демонстрируют нелинейную связь между количеством осадков и ростом растительности, где более высокие уровни осадков, в определенном диапазоне, приводят к более высокой плотности растительности, но также вызывают переходы между различными типами паттернов.
- Экологический стресс/конкуренция за ресурсы ($p$): Рисунок 6 показывает, что по мере медленного увеличения параметра $p$ (связанного с экологическим стрессом или конкуренцией за ресурсы) растительные паттерны, сохраняя свою полосовидную структуру, становятся уже и более концентрированными. Это демонстрирует, что увеличение стресса или конкуренции приводит к более компактному и менее обширному распределению растительности.
Таким образом, численные симуляции не только подтвердили теоретические предсказания устойчивости и неустойчивости, но и предоставили богатое визуальное полотно динамического формирования паттернов. Эти убедительные доказательства, полученные путем систематического варьирования ключевых параметров, окончательно доказали, что основной механизм модели, включающий двойные члены насыщения, точно улавливает сложную экогидрологическую обратную связь, наблюдаемую в регионах, подверженных опустыниванию.
Ограничения и будущие направления
Хотя данное исследование представляет собой строгое и глубокое исследование новой модели растительности и воды, важно признать ее присущие ограничения и рассмотреть направления для будущего развития.
Одно из непосредственных ограничений связано с увеличением нелинейности, введенным членами двойного насыщения. Как отмечают авторы, это значительно повышает аналитическую сложность, особенно при выводе стационарных решений и проведении анализа бифуркаций. Хотя статья успешно справляется с этими проблемами, вычислительная интенсивность и математическая сложность могут стать барьером для более широкого применения или для включения еще более сложных экологических взаимодействий. Будущие исследования могли бы изучить продвинутые численные методы или подходы машинного обучения для эффективной обработки таких нелинейностей высокого порядка, потенциально обеспечивая более быстрые симуляции и более широкое исследование пространства параметров.
Объем численных симуляций — еще один пункт для обсуждения. Эксперименты проводились на конкретной двумерной пространственной области ($[0,50] \times [0,50]$) и конечного временного интервала ($[0,600]$). Хотя этого достаточно для проверки теоретических утверждений, обобщаемость этих выводов на гораздо более крупные географические масштабы, более длительные экологические временные шкалы или различные граничные условия остается открытым вопросом. Будущая работа могла бы включать масштабирование этих симуляций до регионального или континентального уровня, возможно, путем интеграции с данными географических информационных систем (ГИС), для оценки предсказательной силы модели в реальных, гетерогенных ландшафтах.
Кроме того, модель, хотя и продвинутая, в основном фокусируется на взаимодействиях растительности и воды. Другие критические экологические факторы, влияющие на опустынивание, такие как различные типы почв, топография, давление выпаса скота, режимы пожаров или влияние различных видов растений (например, травы против кустарников), не детализированы явно в основных уравнениях модели. Включение этих элементов могло бы привести к еще более полному и реалистичному представлению динамики опустынивания. Например, как изменились бы наблюдаемые паттерны, если бы водоудерживающая способность почвы варьировалась пространственно, или если бы было введено давление выпаса в качестве динамической переменной? Это могло бы привести к многовидовой модели растительности или модели с пространственно явной экологической неоднородностью.
Начальные условия для симуляций включали "небольшие случайные возмущения". Было бы ценно исследовать поведение системы при более крупных или более структурированных возмущениях, которые могли бы лучше имитировать реальные нарушения, такие как экстремальные погодные явления или человеческое вмешательство. Это могло бы выявить различные переходные динамики или альтернативные стабильные состояния, не охваченные малыми возмущениями.
Заглядывая вперед, из этих выводов возникают несколько тем для обсуждения:
- Предиктивное моделирование и системы раннего предупреждения: Учитывая способность модели генерировать разнообразные пространственные паттерны, отражающие наблюдаемую динамику растительности, как это можно использовать для разработки предиктивных инструментов для опустынивания? Могут ли конкретные переходы паттернов (например, от пятен к полосам к пробелам) служить ранними предупреждающими индикаторами деградации экосистем в полузасушливых регионах? Это потребует обширной калибровки с данными долгосрочного экологического мониторинга.
- Оптимизация управления ресурсами: Выводы о том, как диффузия воды ($d_1$) и осадки ($s$) влияют на паттерны растительности, имеют решающее значение. Как эти выводы могут информировать стратегии устойчивого распределения воды в регионах с дефицитом воды? Например, понимание критических значений $d_1$, которые приводят к разреженным пятнам, может помочь менеджерам земель в реализации мер по улучшению удержания влаги в почве или снижению стока, тем самым способствуя более устойчивым структурам растительности.
- Сценарии изменения климата: Помимо простых изменений интенсивности осадков, как модель отреагирует на более сложные прогнозы изменения климата, такие как увеличение частоты и интенсивности засух, сдвиги в сезонных режимах осадков или повышение температуры, влияющее на скорость испарения? Интеграция выходных данных климатических моделей в эту структуру может дать ценное представление о будущих рисках опустынивания.
- Междисциплинарное сотрудничество: Чтобы действительно развить эти выводы, необходимо тесное сотрудничество с полевыми экологами, гидрологами и специалистами по дистанционному зондированию. Проверка предсказаний модели на эмпирических данных из различных регионов, подверженных опустыниванию, укрепит ее экологическую релевантность и практическую применимость. Это также может включать использование данных дистанционного зондирования для вывода паттернов растительности и доступности воды, которые затем могут быть использованы для калибровки и уточнения параметров модели.
- Стохастичность и неопределенность: В статье упоминаются предыдущие работы, включающие стохастичность [23]. Как модель двойного насыщения будет вести себя при стохастических экологических флуктуациях, отражающих присущую неопределенность и изменчивость в естественных системах? Это могло бы дать более надежное понимание устойчивости растительности в непредсказуемых условиях.
- Анализ бифуркаций в высших размерностях: Хотя статья углубляется в локальный и глобальный анализ бифуркаций, сложность модели означает, что полный аналитический анализ всех бифуркационных явлений в пространственных настройках более высокой размерности остается значительной проблемой. Дальнейшие теоретические достижения в этой области могли бы раскрыть более глубокое понимание богатой динамики модели.
Решая эти ограничения и исследуя эти будущие направления, основополагающая работа, представленная в данной статье, может быть значительно расширена, способствуя разработке более эффективных стратегий борьбы с опустыниванием и содействия экологической устойчивости.
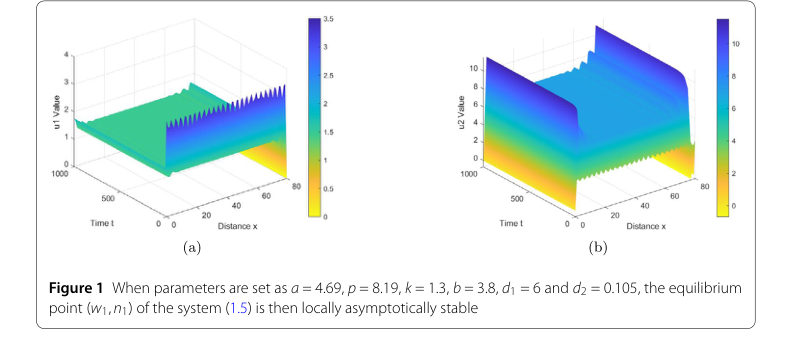 Figure 1. When parameters are set as a = 4.69, p = 8.19, k = 1.3, b = 3.8, d1 = 6 and d2 = 0.105, the equilibrium point (w1,n1) of the system (1.5) is then locally asymptotically stable
Figure 1. When parameters are set as a = 4.69, p = 8.19, k = 1.3, b = 3.8, d1 = 6 and d2 = 0.105, the equilibrium point (w1,n1) of the system (1.5) is then locally asymptotically stable
 Figure 2. When parameters are set as a = 4.69, p = 8.19, k = 1.3, b = 3.8, d1 = 6 and d2 = 0.05, the equilibrium point (w1,n1) of system (1.5) is Turing unstable
Figure 2. When parameters are set as a = 4.69, p = 8.19, k = 1.3, b = 3.8, d1 = 6 and d2 = 0.05, the equilibrium point (w1,n1) of system (1.5) is Turing unstable
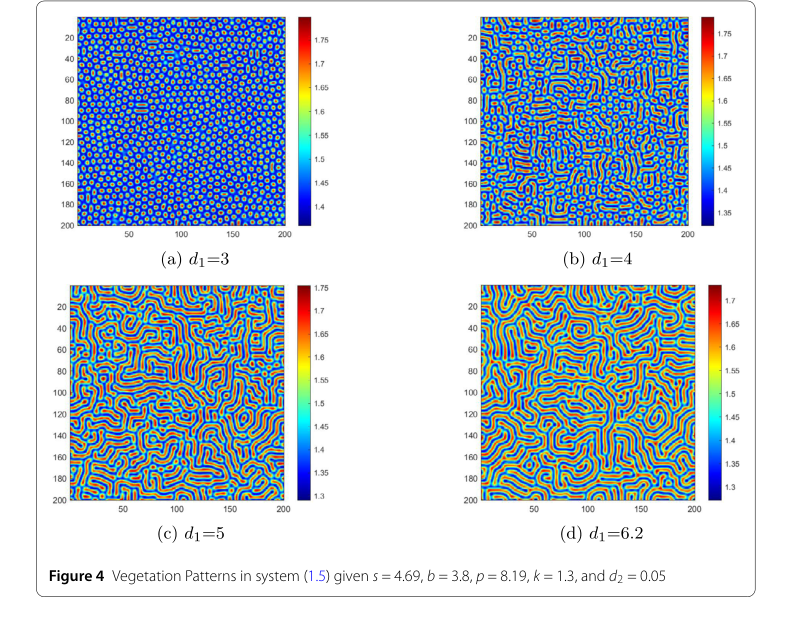 Figure 4. Vegetation Patterns in system (1.5) given s = 4.69, b = 3.8, p = 8.19, k = 1.3, and d2 = 0.05
Figure 4. Vegetation Patterns in system (1.5) given s = 4.69, b = 3.8, p = 8.19, k = 1.3, and d2 = 0.05