植被-水分模型双饱和转化项动力学行为分析
In this paper, we propose a vegetation-water system incorporating double saturation transformation terms, which more vividly depicts the mutual influence and transformation relationship between vegetation and water.
背景与学术渊源
起源与学术渊源
土地荒漠化问题是干旱、半干旱和半湿润地区普遍存在的关键环境问题,构成了本研究的精确起点。全球范围内,由于自然因素和人类活动的加速,干旱、半干旱和半湿润地区荒漠化现象日益严重,导致土壤侵蚀、植被衰退和生态系统稳定性下降。数学生态学领域一直致力于模拟这些复杂的相互作用。
历史上,Klausmeier [8] 提出了一个基础的植被-水分系统(模型1.1),该模型考虑了空间扩散和传输。这一模型为后续研究奠定了基础。例如,Sherratt等人 [9] 集成了脉冲框架来研究间歇性降雨,Consolo等人 [10] 使用广义双曲Klausmeier模型分析了Turing模式。Kealy等人 [11] 通过移除对流项并使用线性扩散简化了Klausmeier模型(得到模型1.2),使其更适用于平坦地形。Sun等人 [12]、Zhang等人 [13] 和 Gou等人 [14] 对这一简化模型进行了进一步研究,以表征稳定构型、分岔参数以及参数对植被分布的影响。随后,Li等人 [29] 在系统中引入了水分吸收饱和效应(模型1.3),认识到植物通过生理调节吸收水分,并在一定的生物量阈值下达到最大吸收能力。
这些先前方法(包括具有单一饱和项的模型1.3)的根本局限性或“痛点”在于,它们无法完全捕捉植被与水分之间双向的饱和相互作用。虽然模型1.3考虑了植被饱和吸水,但并未明确模拟水本身可用性的饱和特性。这一遗漏导致早期模型对水分和植被之间相互影响和转化关系的描绘不够生动和生物学上忠实,尤其是在易荒漠化的地区,水和植被可能同时成为限制因素。这一差距促使作者开发了当前论文的模型,该模型包含一个“双饱和转化项”,以解决这种关键的双向反馈问题,从而在增加分析复杂性的同时,提高了模型的生物学保真度。
直观领域术语
- 土地荒漠化 (Land Desertification): 想象一片曾经繁茂的花园逐渐变成贫瘠的沙地。当土壤失去支持植物生命的能力时,通常是由于自然干旱和过度放牧或森林砍伐等人类活动共同作用,就会发生这种情况。就像花园在慢慢“干涸”并变成沙漠一样。
- Turing不稳定性 (Turing Instability): 想象一下平坦、颜色均匀的蛋糕糊。如果你加入某些成分(例如,反应中的特定化学物质或生物系统中的扩散速率),并让它们相互作用,蛋糕糊可能会自发地形成独特的、重复的图案——如条纹、斑点或迷宫——在凝固过程中。Turing不稳定性描述了这种均匀状态变得不稳定并产生复杂、有组织的图案的现象。
- 双饱和转化项 (Double Saturation Transformation Term): 考虑一株口渴的植物(植被)试图从水源中饮水。首先,无论有多少水可用,植物本身只能吸收一定量的水(植被饱和)。其次,如果土壤中最初的水很少,即使植物能够吸收更多水,它也无法吸收多少(水分饱和)。模型中的这个“双饱和”项同时考虑了这两种限制,使得相互作用更加现实。
- 分岔现象 (Bifurcation Phenomena): 想象一条河流以单一、稳定的水流流淌。如果你逐渐改变一个条件,比如土地的坡度或流水的量,河流可能会突然分成两条不同的水道,或者开始剧烈地蜿蜒,甚至干涸。在数学中,分岔是指系统参数的微小、连续变化导致其行为或其解的数量和类型的突然、定性变化。
符号表
| 符号 | 描述 | 类型 |
|---|---|---|
| $w$ | 无量纲化水密度 | 变量 |
| $n$ | 无量纲化植被生物量 | 变量 |
| $x$ | 无量纲化空间坐标 | 变量 |
| $t$ | 无量纲化时间 | 变量 |
| $s$ | 无量纲化产水量(降雨强度) | 参数 |
| $p$ | 与植被生长/水分转化相关的无量纲化参数 | 参数 |
| $b$ | 无量纲化植被衰减率 | 参数 |
| $k$ | 无量纲化水分饱和阈值 | 参数 |
| $d_1$ | 无量纲化土壤水分扩散系数 | 参数 |
| $d_2$ | 无量纲化植被扩散系数 | 参数 |
问题定义与约束
核心问题表述与困境
本文解决的核心问题是开发一个更具生物学忠实性的数学模型,用于描述易荒漠化地区的植被-水分相互作用。先前的模型,如Klausmeier模型(1.1)及其扩展,如Kealy等人(1.2)和Li等人(1.3)的模型,考虑了植被吸水能力的饱和效应,用类似 $F(N) = \frac{aN^2}{1+bN}$ 的项表示。
输入/当前状态是一系列现有的植被-水分模型,这些模型虽然具有开创性,但未能充分捕捉植被与水分之间双向的饱和相互作用。具体而言,它们缺乏一种机制来考虑水本身可用性的饱和性,导致对生态水文反馈的描绘不够现实。
输出/目标状态是一个新的植被-水分系统,特别是模型(1.4)及其无量纲形式(1.5),它包含了双饱和转化项。这个新模型旨在更准确、更生动地描述植被与水分之间的相互影响和转化关系,从而提高其在描述易荒漠化地区水分-植被相互作用时的生物学保真度。最终目标是在这些改进的条件下,能够对全局稳定性、Turing不稳定性以及各种分岔现象等动力学行为进行更深入、更现实的分析。
本文试图弥合的确切缺失环节或数学鸿沟在于水分的饱和动力学。虽然先前的模型考虑了植被吸水的饱和性($F(N)$),但并未明确模拟水分本身的饱和性。本文引入了一个水分饱和函数 $G(W) = \frac{W}{W+K}$(其中$W$是水密度,$K$是饱和常数)。关键的创新在于组合的双饱和项 $RG(W) \cdot F(N) \cdot N$(或其无量纲等价形式 $\frac{pwn^2}{(w+k)(n+1)}$)。该项精确地刻画了植被饱和吸水能力和水分自身饱和特性协同限制,从而更准确地阐明了非线性生态水文反馈。
作者明确指出的、困扰先前研究人员的痛苦权衡或困境是生物学现实性与分析可处理性之间的平衡。论文指出:“水分饱和函数G(W)的引入极大地提高了模型的生物学保真度,使其能够更真实地描述易荒漠化地区的水分-植被相互作用。然而,由于引入了高阶非线性,这也增加了分析的复杂性,特别是在推导稳态解和进行分岔分析方面。”这意味着,实现更准确的生物学表示是以数学求解和分析系统难度显著增加为代价的。
约束与失效模式
准确模拟易荒漠化地区植被-水分动态的问题极其困难,因为作者们遇到了几个严峻的现实障碍:
-
物理和生态约束:
- 非负解: 对于任何生物模型,水($w$)和植被($n$)的密度都必须保持非负。系统(1.5)服从非负初始数据 $(w_0(x,0), n_0(x,0)) \ge (0,0)$,这是物理现实性的基本要求。未能保持非负性将使模型在生物学上毫无意义。
- 有界域和零通量边界条件: 分析是在有界空间域 $\Omega$ 上进行的,并具有光滑边界,服从零通量(Neumann)边界条件($\frac{\partial w}{\partial \nu} = \frac{\partial n}{\partial \nu} = 0$ 在 $\partial \Omega$ 上)。这些条件代表了一个封闭系统,没有水或植被穿过边界,这对于许多生态斑块来说是一个现实的假设,但增加了偏微分方程(PDEs)的复杂性。
- 饱和现象: 问题的核心在于准确捕捉植被吸水和水本身可用性同时饱和的现象。这是必须嵌入模型中的复杂生理和环境现实。
- 环境因素: 模型必须隐式或显式地考虑影响植被分布和水动力学的各种自然因素(降雨、风速、光照)和人类活动(畜牧业)。
-
计算和数学约束:
- 高阶非线性: 双饱和项 $G(W) \cdot F(N)$ 的引入显著增加了系统(1.4)及其无量纲形式(1.5)的非线性度。这使得稳态解的推导、稳定性分析和分岔分析比线性或低阶非线性系统更加困难。标准分析技术通常难以处理如此高的非线性度。
- 稳态解的分析复杂性: 对于高度非线性的反应-扩散系统,找到显式的稳态解通常非常困难。本文依赖于严格的数学推导、先验估计和定性分析,这些方法计算量大,需要高级数学工具。
- 分岔分析挑战: 分析分岔现象,特别是与简单和双重特征值相关的分岔,需要复杂的L-S降方法和Crandall-Rabinowitz定理等方法。这些方法很复杂,并且随着非线性度的增加,其应用变得更加棘手。本文指出,在某些条件下(例如,$j \neq 2i$ 且 $i \neq 2j$),无法应用隐函数定理来建立解的存在性,这表明标准分析方法存在失效模式。
- Turing不稳定性条件: 确定Turing不稳定性(导致模式形成)发生的条件,涉及对线性化系统的复杂特征值分析,这高度依赖于扩散系数和其他参数。
- 参数空间探索: 模型涉及多个参数(例如,$s, p, b, k, d_1, d_2$)。彻底研究每个参数对系统动力学行为和模式形成的影响,需要广泛的理论分析和数值模拟,使得问题空间庞大且难以完全探索。本文的数值模拟对于跨越这个复杂参数景观验证理论预测至关重要。
为什么选择这种方法
选择的必然性
本文作者面临现有植被-水分模型的一个根本性限制:它们未能充分捕捉易荒漠化地区植被与水分之间复杂的双向饱和效应。传统模型,如Klausmeier模型(系统1.1)及其扩展(如Li等人提出的系统1.3),主要包含一个单一的饱和项,通常描述植被的吸水能力如何随着生物量的增加而饱和,用类似 $f(N) = \frac{aN^2}{1+bN}$ 的函数表示。
然而,作者们意识到这还不够。这种认识的确切时刻在第3页得到了阐述,他们指出:“为了更好地捕捉易荒漠化地区植被与水分之间的双向饱和相互作用,我们通过引入双饱和转化机制进一步完善了系统(1.3)。”这突显了问题不仅在于植被吸水,还在于水分本身的饱和特性以及它如何同时限制植被生长。如果不考虑两者,模型就缺少了生态拼图的关键一块。因此,包含双饱和项的模型不仅仅是改进,而是准确表示这些复杂相互限制的唯一可行解决方案。这是一个明显的不足。
比较优势
这种方法的定性优越性直接源于其增强的生物学保真度,而不仅仅是计算意义上的性能指标改进。引入新的非线性耦合项 $RG(W) \cdot F(N) \cdot N$ 所产生的“双饱和转化项”是关键的结构优势。该项如第3页所述,“刻画了吸水动力学中水分和植被的同时限制”。
先前模型仅考虑单一饱和,无法完全表示这种协同限制。引入水分饱和函数 $G(W)$ 和植被吸收率 $F(N)$ 使模型能够“更真实地描述易荒漠化地区的水分-植被相互作用”并“更准确地阐明生态水文反馈的非线性机制”。这意味着模型不仅仅是更好地拟合数据;它在结构上被设计成更忠实地反映现实世界的生态过程。它提供了对这两种关键成分在压力下如何相互作用的更深入、更细致的理解,这比先前的黄金标准有了显著的定性飞跃。本文没有讨论处理高维噪声或降低内存复杂度等问题,因为它专注于基础的生态现象数学建模。
与约束的契合
坦率地说,我不太确定第2步中定义的具体约束,因为该部分未提供。然而,根据论文中提出的问题定义和动机,我们可以推断出几种该选择方法完美解决的严峻要求。核心问题围绕着准确模拟“易荒漠化地区”(摘要,第3页)的植被-水分相互作用,这意味着需要高度保真地捕捉环境压力下的关键反馈机制。
问题要求与解决方案的独特性质之间的“结合”体现在几个方面:
1. 真实地描绘相互影响: 问题要求一个模型能够“更生动地描绘植被与水分之间的相互影响和转化关系”(摘要)。双饱和项 $G(W)$ 和 $F(N)$ 通过明确建模水分可用性和植被吸水能力如何同时限制彼此,直接满足了这一要求。
2. 双向饱和相互作用: 作者们明确寻求“更好地捕捉植被与水分之间的双向饱和相互作用”(第3页)。新颖的双饱和项 $RG(W) \cdot F(N) \cdot N$ 正是为了此目的而设计的,确保模型考虑了相互作用双方的饱和性,这是先前模型中的一个关键缺失环节。
3. 阐明非线性机制: 荒漠化涉及复杂的非线性生态水文反馈。所选方法通过双饱和项引入的高阶非线性,能够“更准确地阐明易荒漠化地区生态水文反馈的非线性机制”(第3页)。这与理解驱动这些脆弱环境中模式形成和生态系统稳定性的复杂动力学的需求完美契合。
替代方案的拒绝
本文作者拒绝替代方案的理由主要是隐含的,并且侧重于先前数学模型的局限性,而不是完全不同的方法学范式,如GANs或Transformers。作者们在反应-扩散模型的基础上进行研究,从Klausmeier系统(1.1)及其后续扩展(例如,Kealy等人系统(1.2)、Li等人系统(1.3))开始。
拒绝这些早期模型作为充分模型的根本原因是它们无法捕捉“植被与水分之间的双向饱和相互作用”(第3页)。虽然这些模型包含了单一饱和项(例如,植被吸水的 $f(N) = \frac{aN^2}{1+bN}$),但它们缺乏相应的水分饱和项 $G(W)$。作者们发现,“传统”方法(在此语境下指单一饱和植被-水分模型)“不足以解决此特定问题”,因为它们无法“同时捕捉植被吸水能力和水分可用性本身的饱和效应”(第4页)。这一遗漏导致生态水文反馈的描绘不够现实。因此,具有双饱和项的新模型被引入,作为克服同一建模框架内其前身这些特定缺点的必要改进。本文没有深入探讨为什么GANs或Diffusion模型等机器学习模型会失败,因为它们的操作层面不同,并且与本文所采取的机械微分方程建模方法不直接可比。
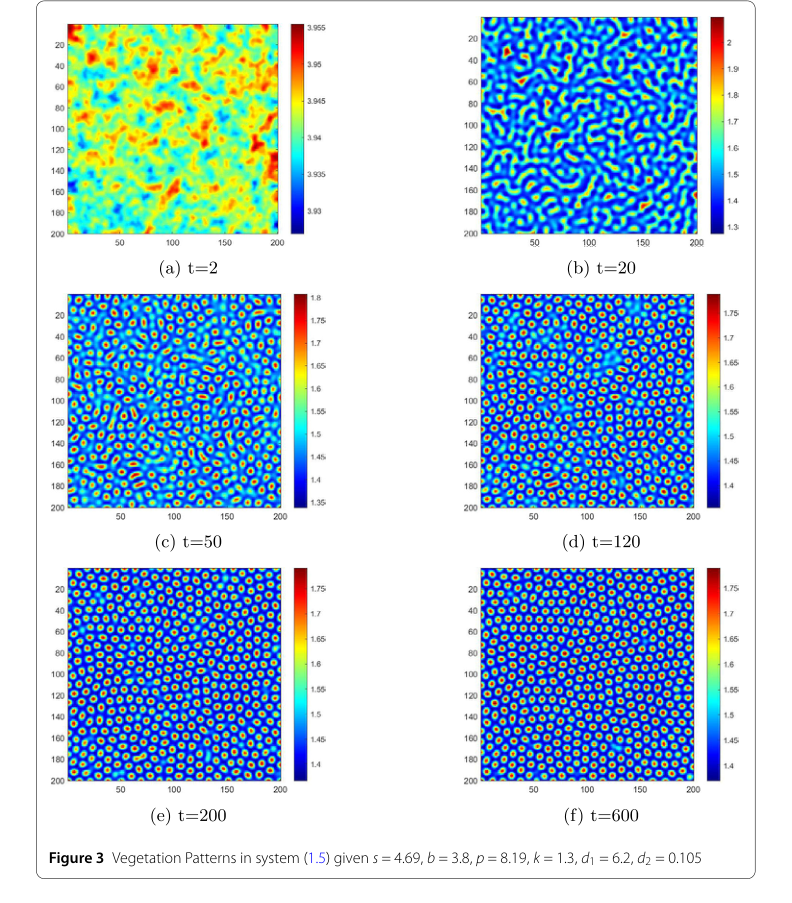 Figure 3. Vegetation Patterns in system (1.5) given s = 4.69, b = 3.8, p = 8.19, k = 1.3, d1 = 6.2, d2 = 0.105
Figure 3. Vegetation Patterns in system (1.5) given s = 4.69, b = 3.8, p = 8.19, k = 1.3, d1 = 6.2, d2 = 0.105
数学与逻辑机制
主方程
本文数学引擎的核心是无量纲化的反应-扩散系统,它描述了植被生物量和水密度之间的动力学相互作用。该系统通过引入双饱和转化项并进行尺度化后得到,表示为:
$$ \begin{cases} \frac{\partial w}{\partial t} = s - w - \frac{pwn^2}{(w+k)(n+1)} + d_1\Delta w, & x \in \Omega, t > 0, \\ \frac{\partial n}{\partial t} = \frac{pwn^2}{(w+k)(n+1)} - bn + d_2\Delta n, & x \in \Omega, t > 0, \\ \frac{\partial w}{\partial \nu} = \frac{\partial n}{\partial \nu} = 0, & x \in \partial\Omega, t > 0, \\ (w_0(x, 0), n_0(x, 0)) \geq, \neq (0,0), & x \in \Omega. \end{cases} $$
各项剖析
让我们逐一剖析这个主方程,以理解其组成部分及其作用:
-
$w$:
- 数学定义: 一个因变量,表示无量纲化的水密度。它是一个标量场,意味着其值可以随空间和时间变化。
- 物理/逻辑作用: 该项量化了在任何给定位置 $x$ 和时间 $t$ 生态系统中可用水的量。
- 为何使用: 它是水的主要状态变量。
-
$n$:
- 数学定义: 一个因变量,表示无量纲化的植被生物量密度。与 $w$ 类似,它是一个标量场。
- 物理/逻辑作用: 该项量化了在任何给定位置 $x$ 和时间 $t$ 存在的植被生物量。
- 为何使用: 它是植被的主要状态变量。
-
$t$:
- 数学定义: 代表时间的自变量。
- 物理/逻辑作用: 这是系统演变的时间维度。
- 为何使用: 描述 $w$ 和 $n$ 如何随时间变化。
-
$x \in \Omega, t > 0$:
- 数学定义: 指定偏微分方程(PDEs)的域。$\Omega$ 是一个有界空间区域,$t > 0$ 表示我们关注的是初始时刻之后的系统演变。
- 物理/逻辑作用: 定义了模型适用的物理空间和时间间隔。
- 为何使用: 为空间和时间动力学设定背景。
-
$\frac{\partial w}{\partial t}$:
- 数学定义: 水密度 $w$ 对时间 $t$ 的偏导数。
- 物理/逻辑作用: 这表示在空间特定点处水密度瞬时变化率。正值表示水在增加,负值表示在减少。
- 为何使用: 模拟水的时变演化。
-
$\frac{\partial n}{\partial t}$:
- 数学定义: 植被生物量密度 $n$ 对时间 $t$ 的偏导数。
- 物理/逻辑作用: 这表示空间特定点处植被生物量瞬时变化率。
- 为何使用: 模拟植被的时变演化。
-
$s$:
- 数学定义: 一个正的常数参数。
- 物理/逻辑作用: 代表进入系统的恒定水量输入,类似于降雨或降水。它是水的一个源项。
- 为何使用: 模拟连续的外部水源供应。它是一个加法项,因为它直接贡献于水库。
-
$-w$:
- 数学定义: 一个线性项,水密度 $w$ 的负值。
- 物理/逻辑作用: 代表蒸发导致的水损失率。在原始的量纲模型中,这是 $LW$,其中 $L$ 是蒸发率。无量纲化后,$L$ 被隐式吸收或设为1。它是水的一个汇项。
- 为何使用: 模拟与水量成比例的水损失。它是一个减法项,因为水从系统中移除。
-
$-\frac{pwn^2}{(w+k)(n+1)}$(在第一个方程中):
- 数学定义: 一个涉及 $p, w, n, k$ 的复杂非线性项。
- 物理/逻辑作用: 这是关键的“双饱和转化项”,表示植被吸收水分的速率。$p$ 是转化效率。$n^2$ 表明吸水率随植被密度增加而增加得比线性更快,可能由于协同效应或表面积增加。分母 $(w+k)$ 和 $(n+1)$ 引入了饱和效应:当水 $w$ 或植被 $n$ 非常高时,吸水速率不会无限增加,而是趋于最大值。该项从水方程中减去,因为水被植被消耗。
- 为何使用: 捕捉植被通过水分吸收的现实生物学现象,包括水可用性和植被吸收能力饱和效应。乘法组合了 $p, w, n^2$ 的影响。除以 $(w+k)$ 和 $(n+1)$ 产生了饱和效应,即速率在高值时趋于平缓,这在生物学中是模拟有限资源或能力的常见方式。
-
$+d_1\Delta w$:
- 数学定义: 一个扩散项,其中 $d_1$ 是正常数,$\Delta$ 是拉普拉斯算子($\Delta w = \frac{\partial^2 w}{\partial x_1^2} + \frac{\partial^2 w}{\partial x_2^2} + \dots$)。
- 物理/逻辑作用: 代表由于扩散(例如,土壤渗透、侧向流动)导致的水的空间扩散。水倾向于从高浓度区域流向低浓度区域,平滑空间差异。正号表示这种平滑效应。
- 为何使用: 模拟水在空间中自然分散的趋势。它是一个加法项,因为它代表一个净通量,该通量可以根据水剖面的曲率增加或减少局部水密度。
-
$\frac{pwn^2}{(w+k)(n+1)}$(在第二个方程中):
- 数学定义: 与第一个方程中的非线性项相同。
- 物理/逻辑作用: 该项表示植被通过转化吸收的水来生长成生物量的速率。它是植被的一个源项。
- 为何使用: 将水分吸收直接与植被生长联系起来。它是一个加法项,因为这个过程增加了植被生物量。
-
$-bn$:
- 数学定义: 一个线性项,是 $b$ 和 $n$ 的乘积。$b$ 是一个正常数。
- 物理/逻辑作用: 代表植被生物量的自然死亡或损失率。它是植被的一个汇项。
- 为何使用: 模拟植被的自然衰减或死亡。它是一个减法项,因为植被从系统中移除。
-
$+d_2\Delta n$:
- 数学定义: 一个扩散项,其中 $d_2$ 是正常数,$\Delta$ 是应用于 $n$ 的拉普拉斯算子。
- 物理/逻辑作用: 代表植被的空间扩散(例如,通过无性繁殖、种子传播)。植被倾向于从高密度区域扩散到低密度区域。
- 为何使用: 模拟植被在空间中自然分散的趋势。与 $d_1\Delta w$ 原因相同,它是一个加法项。
-
$\frac{\partial w}{\partial \nu} = \frac{\partial n}{\partial \nu} = 0, x \in \partial\Omega, t > 0$:
- 数学定义: 诺依曼边界条件,其中 $\frac{\partial}{\partial \nu}$ 表示法向导数(垂直于边界的变化率)。
- 物理/逻辑作用: 这些条件在空间域边界施加了“零通量”或“无流动”约束。这意味着没有水或植被生物量进入或离开系统边界,有效地使系统成为一个封闭单元。
- 为何使用: 定义一个封闭系统,其中所有相互作用都发生在指定域内,防止外部影响影响边界动力学。
-
$(w_0(x, 0), n_0(x, 0)) \geq, \neq (0,0), x \in \Omega$:
- 数学定义: 水和植被密度的初始条件。它们是非负的,并且不处处为零。
- 物理/逻辑作用: 指定初始时间 $t=0$ 时水和植被生物量的起始空间分布。非零条件确保了动力学展开的初始“种子”。
- 为何使用: 为系统演变模拟或分析提供起点。
逐步流程
想象一下我们生态域内特定位置共存的一小块抽象的水和一小片植被。让我们追踪它们在数学引擎中的旅程:
- 水分输入: 首先,水块接收恒定的稳定水量输入 $s$,类似于降雨。这是它的主要输入。
- 水分损失(蒸发): 同时,水块中的一部分水通过蒸发损失到环境中,由 $-w$ 项表示。水越多,蒸发越多。
- 植被的渴求(从水的角度): 接下来,该位置的植被开始从水块中吸收水分。这种吸收是一个复杂的过程:它由植被密度 ($n^2$) 和效率因子 ($p$) 驱动,但也是有限制的。当水块变得非常湿润(高 $w$)或植被密度非常高(高 $n$)时,吸收速率不会无限增加;它会“饱和”或趋于平缓,这得益于分母中的 $(w+k)$ 和 $(n+1)$ 项。吸收的水从水块中移除。
- 水分扩散: 在这些局部相互作用之后,水块中的水不会停留在原地。它会扩散到邻近区域,同时邻近区域的水也会扩散到这个水块中。这个过程由扩散系数 $d_1$ 和拉普拉斯算子 $\Delta w$ 控制,就像一个平滑机制,倾向于使空间中的水浓度均衡。
- 植被生长(从植被的角度): 现在,切换到植被。从水块中吸收的($\frac{pwn^2}{(w+k)(n+1)}$ 项)正是这些水被转化为新的植被生物量。这是植被的主要生长机制,直接将其扩张与水分可用性和自身密度联系起来。新的生物量被添加到植被中。
- 植被损失(死亡): 然而,植被也面临自然损失。一部分植被生物量,与其当前密度成比例 ($bn$),会死亡或因其他因素而损失。这从植被中减去。
- 植被扩散: 类似于水,植被生物量也会在空间上扩散。在扩散系数 $d_2$ 和拉普拉斯算子 $\Delta n$ 的驱动下,植被会扩散到密度较低的区域,新的植被也会从邻居扩散到这个区域。
- 边界约束: 在整个生态域的最外围,施加了“零通量”条件。这意味着没有水或植被生物量可以穿过这些边界进出系统,有效地使系统成为一个自给自足的单元。
- 连续演化: 这些步骤不是顺序发生的,而是在域内每个点上连续且同时发生的。随着时间的推移,从水和植被的初始分布开始,这种局部反应(输入、蒸发、吸收、生长、损失)和空间扩散的复杂相互作用导致整个生态系统演化,可能导致稳定的均匀状态或复杂、动态的空间模式。
优化动力学
在这个反应-扩散系统的背景下,“优化动力学”并非指模型从数据中学习或在机器学习中最小化损失函数,而是指系统如何演化并收敛到稳定状态或模式。这里的“学习”是系统内在的、寻找其自然配置的趋势。
-
向平衡点的演化: 系统的主要动力学是向平衡点演化,在这些平衡点处,水和植被的变化率都变为零($\frac{\partial w}{\partial t} = 0$ 且 $\frac{\partial n}{\partial t} = 0$)。这些平衡点代表稳态,在这些稳态下,输入和输出以及反应和扩散过程完美平衡。本文确定了几个这样的点:一个“边界平衡点”(例如,只有水没有植被的状态)和“正平衡点”(同时有水和植被的状态)。
-
稳定性和“损失景观”: “损失景观”的概念可以直观地映射到这些平衡点的稳定性景观。如果一个平衡点是稳定的,系统在受到轻微扰动后会倾向于返回该状态。如果不稳定,即使是微小的扰动也会导致系统远离。这种景观的形状由非线性相互作用和扩散项决定。本文通过检查线性化系统在平衡点周围雅可比矩阵的特征值来使用线性稳定性分析。
- 对于均匀平衡点,如果所有特征值的实部都为负,则它是稳定的。如果任何特征值的实部为正,则它是不稳定的。
-
Turing不稳定性:模式形成: 一个关键的动力学是Turing不稳定性,这是空间模式如何产生的方式。当一个空间均匀的平衡点对均匀扰动是稳定的(意味着它在均匀扰动后会恢复均匀性),但对空间变化的扰动变得不稳定(意味着某些空间波长的扰动会增长)时,就会发生这种情况。
- 扩散系数($d_1$ 和 $d_2$)在这里至关重要。如果它们足够不同,它们就可以在特定空间频率下使均匀状态不稳定,从而导致斑点、条纹或间隙等模式的自发形成。这就像系统“找到”了一个更复杂、空间异质性的“最优解”。本文通过分析线性化系统(现在包含扩散项)的特征值在不同空间波数下的行为来分析这一点。
-
分岔动力学:定性转变: 随着关键参数(如水扩散系数 $d_1$)的变化,系统会发生分岔。这意味着解的定性性质会发生剧烈变化。例如,一个稳定的均匀状态可能会突然产生稳定的空间模式,反之亦然。
- 本文研究了稳态分岔,其中新的平衡解(通常是非均匀模式)从现有解中分支出来。这些分岔的方向(超临界或次临界)决定了新模式是否稳定并平滑出现(超临界)还是不稳定并突然出现(次临界),可能导致滞后现象。这由分岔函数的更高阶导数决定。
-
迭代状态更新(数值模拟): 虽然数学分析提供了关于稳定性和分岔的理论见解,但系统状态随时间的实际迭代更新是通过数值模拟来可视化的。通过离散化空间和时间来计算求解偏微分方程。在每个小时间步长中,基于局部反应项和邻近点的扩散来更新每个空间网格点上的 $w$ 和 $n$ 值。这个迭代过程允许研究人员观察空间模式的出现和演变,确认理论预测并说明系统如何“收敛”到其动态配置。本文的图清晰地展示了这一点,显示了在不同参数设置下,初始随机噪声如何演变成有组织的斑点、条纹或间隙模式。
结果、局限性与结论
实验设计与基线
本研究的理论发现的实验验证是通过对提出的植被-水分系统(1.5)进行数值模拟来完成的。主要目标是严格证明关于稳定性、不稳定性以及模式形成的数学论断。作者通过选择两种不同的参数配置来构建他们的实验,每种配置都旨在与特定的理论预测相符。
对于第一组参数,旨在证明局部渐近稳定性,其值设定为 $s = 4.69$, $p = 8.19$, $k = 1.3$, $b = 3.8$, $d_1 = 6$, 和 $d_2 = 0.105$。这些参数经过专门选择,以满足条件 $z < \dot{z} < z_2$,根据定理2.2,该条件预测了平衡点 $(w_1, n_1)$ 的局部渐近稳定性。
相反,对于第二组参数,旨在说明Turing不稳定性,其值设定为 $s = 4.69$, $p = 8.19$, $k = 1.3$, $b = 3.8$, $d_1 = 6$, 和 $d_2 = 0.05$。该配置选择以满足条件 $d_2\lambda_1 < \frac{b}{n_1+1}$ 和 $d_1 > \bar{d_1}$,根据定理2.5 (2),这些条件应导致平衡点 $(w_1, n_1)$ 的Turing不稳定性状态。
模拟在定义为 $[0,50] \times [0,50]$ 的二维空间区域上进行,并在 $[0,600]$ 秒的时间间隔内演化。这些模拟的初始条件包括在空间均匀系统中对稳态解 $(w_1, n_1)$ 引入小的随机扰动。“受害者”在这个实验设计中不是外部基线模型,而是理论预测本身。核心机制工作的确凿证据是直接的视觉和定量确认,即数值模拟准确地重现了预测的稳定或不稳定行为以及随后的模式形成。
证据证明的内容
数值模拟提供了确凿的证据,证明了核心数学机制,特别是双饱和项和扩散系数的作用,正如理论分析所预测的那样。
-
稳定性与不稳定性验证:
- 局部渐近稳定性: 对于第一组参数,图1图形化地展示了系统的演化。图中的“u1值”(水密度)和“u2值”(植被生物量)显示,尽管存在初始扰动,系统在空间域上随时间迅速收敛到一个均匀、稳定的状态。这直接验证了定理2.2,证实了在特定条件下,平衡点 $(w_1, n_1)$ 确实是局部渐近稳定的。
-
Turing不稳定性: 对于第二组参数,图2鲜明地对比了稳定行为。在这里,“u1值”和“u2值”图清楚地显示了复杂空间模式的出现和持续存在,而不是均匀状态。这种确定的模式形成是Turing不稳定的标志,为定理2.5 (2) 的预测提供了确凿证据。系统在初始扰动后演化成一个结构化的、非均匀的状态,证明了理论上的不稳定性条件被准确识别。
-
动态植被模式形成:
- 图3展示了植被分布随时间的变化,揭示了一个引人入胜的模式演化序列。从随机扰动开始,系统最初表现出不规则、非均匀的暂时模式。随着时间的推移(例如,从 $t=2$ s 到 $t=50$ s),条纹状模式开始出现。最终,到 $t=600$ s,这些模式演化成主要的间隙模式,然后保持不变。从初始无序到复杂稳定模式的动态演进,为模型捕捉现实生态现象的能力提供了令人信服的证据。蓝色(低水分)和红色(高水分)区域的视觉区分进一步阐明了资源的分布。
-
关键生态参数的影响:
- 水扩散系数 ($d_1$): 在图4中,第一个关键因素是水扩散系数 $d_1$。当 $d_1 = 3$ 时,如图4 (a) 所示,出现规则的高密度斑块。在这种条件下,水分扩散缓慢,植被以小而集中的斑块形式生长,类似于自然界的草地斑块。随着 $d_1$ 增加,过渡到图4 (b) 的情况,模式演变成复杂的迷宫状条纹,高水/低植被和低水/高植被区域交替出现。当 $d_1$ 达到5时,条纹拉长,形成复杂的带状和斑块混合体;增强的水分扩散拉伸了植被,使其分布更分散。当 $d_1$ 设定为6.2时,如图4 (d) 所示,出现大而稀疏的斑块。高水区域广泛扩张,将植被推入孤立的小区域,导致植被密度下降。这一序列严格证明了水分迁移率的增加(更高的 $d_1$)会促进植被扩散并降低生物量密度。
-
降雨强度 ($s$): 图5展示了降雨率的影响。最初,随着 $s$ 的增加(例如,$s=3.69$ 到 $s=4$),出现条纹模式。进一步增加 $s$(例如,$s=4.5$ 到 $s=4.69$)导致间隙模式的形成。这些证据证明了降雨与植被生长之间存在非线性关系,在特定范围内,更高的降雨率会导致更高的植被密度,但也会驱动不同模式类型之间的转变。
- 环境压力/资源竞争 ($p$): 图6显示,随着参数 $p$(与环境压力或资源竞争相关)的缓慢升高,植被模式虽然保持其条纹状结构,但变得更窄、更集中。这表明增加的压力或竞争会导致更紧凑、分布范围更小的植被。
总而言之,数值模拟不仅验证了稳定性和不稳定性等理论预测,还提供了丰富的动态模式形成的可视化图景。这些来自系统性改变关键参数的硬证据,明确证明了模型的核心机制,包括双饱和项,能够准确捕捉在易荒漠化环境中观察到的复杂生态水文反馈。
局限性与未来方向
尽管本研究提出了对新型植被-水分模型的严格且富有洞察力的分析,但认识到其固有的局限性并考虑未来发展方向至关重要。
一个直接的局限性源于双饱和项引入的非线性度增加。正如作者所指出的,这显著增加了分析的复杂性,特别是在推导稳态解和进行分岔分析方面。虽然本文成功地应对了这些挑战,但计算强度和数学复杂性可能成为更广泛应用或纳入更复杂生态相互作用的障碍。未来的研究可以探索先进的数值技术或机器学习方法来有效地处理这种高阶非线性,从而可能实现更快的模拟和更广泛的参数空间探索。
数值模拟的范围是另一个讨论点。实验是在特定的二维空间域($[0,50] \times [0,50]$)和有限的时间间隔($[0,600]$)内进行的。虽然足以验证理论论点,但这些发现对于更大地理尺度、更长生态时间尺度或不同边界条件的普遍性仍然是一个悬而未决的问题。未来的工作可以考虑将这些模拟扩展到区域或大陆级别,也许通过与地理信息系统(GIS)数据集成,以评估模型在真实世界异质景观中的预测能力。
此外,该模型虽然先进,但主要侧重于植被-水分相互作用。影响荒漠化的其他关键生态因素,如土壤类型变化、地形、牲畜放牧压力、火灾制度或不同植物物种(例如,草与灌木)的影响,在模型核心方程中并未明确详述。纳入这些要素可以更全面、更现实地描绘荒漠化动态。例如,如果土壤保水能力在空间上变化,或者引入动态变量的放牧压力,观察到的模式会如何变化?这可能导致多物种植被模型或具有空间显式环境异质性的模型。
模拟的初始条件涉及“小的随机扰动”。研究系统在更大或更结构化的扰动下的行为将是有价值的,这可能更好地模拟现实世界的干扰,如极端天气事件或人为干预。这可能会揭示与小扰动不同的瞬态动力学或替代稳定状态。
展望未来,从这些发现中涌现出几个讨论话题:
- 预测建模与预警系统: 鉴于模型能够生成反映观察到的植被动态的各种空间模式,如何利用这一点开发荒漠化预测工具?特定的模式转变(例如,从斑点到条纹到间隙)是否可以作为半干旱地区生态系统退化的早期预警指标?这将需要与长期生态监测数据进行广泛校准。
- 优化资源管理: 关于水扩散 ($d_1$) 和降雨 ($s$) 如何影响植被模式的见解至关重要。这些发现如何为水资源稀缺地区的公平水资源分配策略提供信息?例如,理解导致稀疏斑块的关键 $d_1$ 值可以指导土地管理者采取措施,以增强土壤保水能力或减少径流,从而促进更具韧性的植被结构。
- 气候变化情景的影响: 除了简单的降雨强度变化外,模型将如何应对更复杂的气候变化预测,例如干旱频率和强度的增加、季节性降雨模式的转变或影响蒸发率的温度升高?将气候模型输出整合到此框架中,可以为未来的荒漠化风险提供有价值的见解。
- 跨学科合作: 为了真正发展这些发现,与野外生态学家、水文学家和遥感专家的密切合作至关重要。将模型预测与来自不同易荒漠化地区的实证数据进行验证,将加强其生态相关性和实际适用性。这还可能涉及使用遥感数据来推断植被模式和水可用性,然后可以用于校准和完善模型参数。
- 随机性与不确定性: 本文提到了先前工作中包含随机性的研究[23]。在反映自然系统固有的不确定性和变异性的随机环境波动下,双饱和模型将如何表现?这可以提供对植被在不可预测条件下的韧性的更稳健的理解。
- 高维分岔分析: 虽然本文深入研究了局部和全局分岔分析,但模型的复杂性意味着对高维空间设置中所有分岔现象进行全面分析仍然是一个重大挑战。该领域进一步的理论进展可以解锁对模型丰富动力学更深入的理解。
通过解决这些局限性并探索这些未来方向,本文提出的基础工作可以得到显著扩展,为应对荒漠化和促进生态韧性做出更有效的贡献。
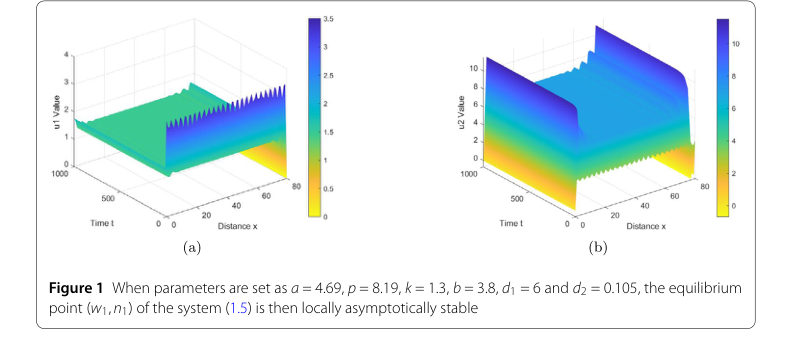 Figure 1. When parameters are set as a = 4.69, p = 8.19, k = 1.3, b = 3.8, d1 = 6 and d2 = 0.105, the equilibrium point (w1,n1) of the system (1.5) is then locally asymptotically stable
Figure 1. When parameters are set as a = 4.69, p = 8.19, k = 1.3, b = 3.8, d1 = 6 and d2 = 0.105, the equilibrium point (w1,n1) of the system (1.5) is then locally asymptotically stable
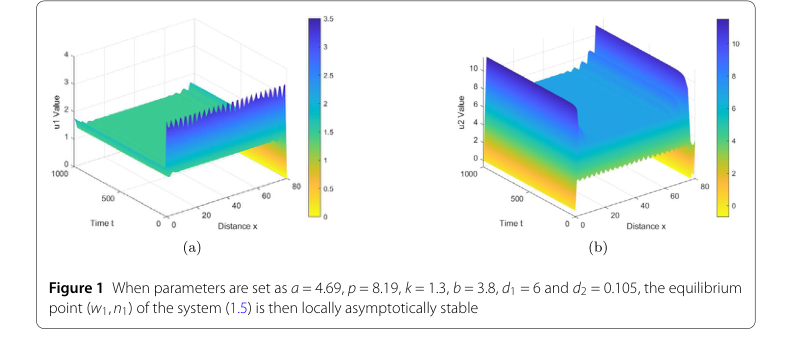 Figure 2. When parameters are set as a = 4.69, p = 8.19, k = 1.3, b = 3.8, d1 = 6 and d2 = 0.05, the equilibrium point (w1,n1) of system (1.5) is Turing unstable
Figure 2. When parameters are set as a = 4.69, p = 8.19, k = 1.3, b = 3.8, d1 = 6 and d2 = 0.05, the equilibrium point (w1,n1) of system (1.5) is Turing unstable
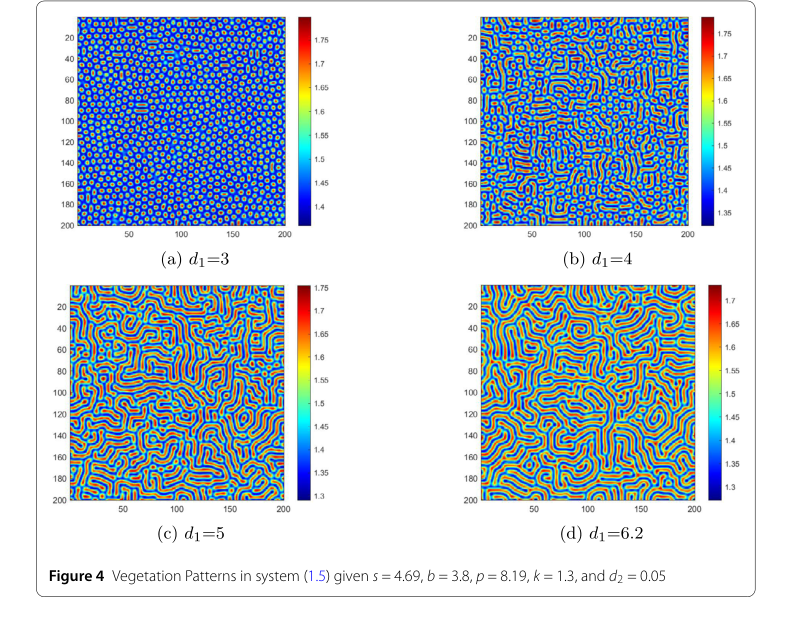 Figure 4. Vegetation Patterns in system (1.5) given s = 4.69, b = 3.8, p = 8.19, k = 1.3, and d2 = 0.05
Figure 4. Vegetation Patterns in system (1.5) given s = 4.69, b = 3.8, p = 8.19, k = 1.3, and d2 = 0.05
与其他领域的同构性
结构骨架
本文提出了一个反应-扩散机制,该机制模拟了两个相互作用量的发展和模式形成,其中包含了非线性饱和动力学,并分析了它们的稳定性和分岔点。